A. La plasticité du génome bactérien

1
Cours n°2
Bactériologie
Professeur Antoine Andremont
Vendredi 3 octobre
8h30
Ronéo typeur : Diana Mourão Balsa
Génétique bactérienne

2
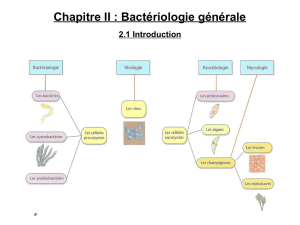
Plan du cours :
Introduction
I. Le génome bactérien, plasticité et stabilité
A. La plasticité du génome bactérien
1. Mise en évidence de la plasticité du génome bactérien par la résistance
bactérienne aux antibiotiques
2. Le monde bactérien, une remarquable adaptation
3. La plasticité du génome bactérien et ses mécanismes
4. Les divers types de bactéries
B. Le génome bactérien
1. Du gène à la protéine chez les bactéries
2. Comparaison schématique de la bactérie et de la cellule eucaryote
3. L’ADN bactérien
C. La stabilité des espèces bactériennes
II. Les mécanismes de plasticité du génome bactérien
A. Les mutations au sein d’un génome
1. Rappel concernant le mécanisme des mutations
2. Expériences démontrant que les mutations surviennent de façon spontanée
3. Un exemple « d’application » des mutations
4. La gravité des mutations en clinique
B. Le transfert de gènes entre les bactéries
1. La transformation
a) Définition
b) Expériences
c) La transformation avec des fragments d’ADN et avec des
plasmides

3
d) Applications et conséquences de la transformation
2. La transduction
a) Définition
b) Rappel : le cycle de vie des bactériophages lytiques
c) La transduction généralisée
d) La transduction spécialisée
e) Une application de la transduction : les cosmides
3. La conjugaison
a) Définition et propriétés
b) Découverte de la conjugaison
c) Les étapes de la conjugaison
d) Le contact cellulaire, rôle du pilus F
e) Conjugaison chez les streptocoques
f) Rôle des plasmides de résistance dans l’évolution de la
résistance
g) Les plasmides et leur application en biotechnologie
C. Les éléments génétiques mobiles à l’intérieur de la bactérie
Conclusion
Tout d’abord ce qui est en italique correspond à ce qui a été dit en cours : commentaires sur
schémas, explications complémentaires au texte.
J’ai enlevé certaines diapos dont le prof ne s’est pas servi, en revanche j’ai jugé utile d’en
laisser certaines.
Bonne année à tous.

4
Introduction : Pourquoi la génétique bactérienne est-elle importante pour
la pratique médicale ?
La résistance des bactéries aux antibiotiques
C’est un problème de santé publique rencontré dans la pratique médicale
La virulence
Du fait que certaines bactéries sont responsables d’infections très dramatiques qui ont
influencé l’évolution, on a l’habitude de voir les bactéries comme étant nos « ennemies ».
En fait la virulence est un moment dans l’évolution des bactéries qui explique que
certaines maladies disparaissent mais aussi que d’autres émergent de façon régulière.
Les biotechnologies
A chaque fois qu’on utilise les produits des biotechnologies on fait appel sans le savoir à
la génétique bactérienne. Aussi le progrès des connaissances dans ce domaine permettent
l’essor des biotechnologies avec l’apparition de nouveaux produits.
I. Le génome bactérien, plasticité et stabilité
A. La plasticité du génome bactérien
1. Mise en évidence de la plasticité du génome bactérien par la résistance
bactérienne aux antibiotiques
La résistance bactérienne aux antibiotiques suit le développement des nouvelles molécules.
Il y a peu (voire pas) d’exemples d’absence de résistance à un antibiotique donné.
Ce qui constitue un apparent paradoxe car de nombreuses molécules sont disponibles.
La réponse est la plasticité du génome bactérien
Ces cinquante dernières années on a observé un développement considérable des antibiotiques, à
chaque fois plus puissants et à spectres plus larges. Mais finalement à chaque fois les bactéries ont su
développer des mécanismes de résistance. L’explication de ce phénomène est la plasticité du génome
bactérien grâce à laquelle ils ont une énorme capacité à s’adapter à l’environnement qui les entoure.
Courbe représentant sur 10 ans l’évolution de la
résistance d’E.Coli à l’amoxicilline.
Constat : cette résistance a augmenté de 15 points
en 10 ans.

5
2. Le monde bactérien, une remarquable adaptation
• L’âge des bactéries : 3.5 milliards d’années
• Les premiers hommes : 3.5 millions d’années (Mille fois moins longtemps)
• Bactéries hautement adaptables (« tous terrains »)
– Plasticité du génome (éléments mobiles)
– Nombre (des milliers d’espèces)
– Lieux (elles sont présentes partout à la surface du globe)
L’âge de la Terre est d’environ 4 milliards d’années ; les bactéries ayant un âge de 3,5 milliards
d’années : elles sont apparues très tôt. Elles se sont donc adaptées à tous les changements
d’environnement qu’elles ont connu depuis 3,5 milliards d’années, colonisé toutes formes d’existence
(dont les êtres humains) et se sont répandues partout à la surface de la Terre. Comment cela est-il
possible ? Encore une fois cette remarquable capacité d’adaptation des bactéries est due à la plasticité
du génome bactérien.
Une chose impressionnante : nous avons dans notre corps plus de bactéries que de cellules eucaryotes.
3. La plasticité du génome bactérien et ses mécanismes
→ « C’est l’imagination des bactéries pour trouver des solutions à des pressions de sélection
environnementales. »
Il y a globalement 2 mécanismes de plasticité du génome : les mutations et l’acquisition de
nouveaux gènes.
• Les mutations : survenue constante, simplicité et rapidité pour modifier une fonction mais système
simple (pas de solution complexe), transfert vertical uniquement
• L’acquisition de « nouveaux » gènes : survenue rare, long pour « fabriquer les premiers éléments
», mais permet des solutions complexes et multiples, transfert vertical et horizontal.
Mutations : changements singuliers de paires de base. Effet immédiat mais au même temps ce sont
des choses ponctuelles qui engendrent des réponses « oui/non » comme par exemple la perte d’une
fonction. Ce système ne permet donc pas d’adapter finement la réponse bactérienne à un changement
subtil.
La transmission verticale c’est la transmission de matériel génétique entre une bactérie mère et une
bactérie fille. Finalement le système des mutations ne permet pas à lui seul d’expliquer l’évolution
car la vitesse et la rusticité de ce système ne permettent pas de comprendre comment en un temps
long mais fini on a pu passer de bactéries à des êtres aussi complexes que les hommes.
Acquisition de nouveaux gènes : Les bactéries en plusieurs millions d’années dans un écosystème
particulier finissent par fabriquer une fonction complexe avec tout un système de régulation comme
par exemple le fait de sentir de façon très précise un composé de leur environnement et de réagir en
fonction de la concentration de celui-ci.
Une fois que ce système a été développé par une bactérie d’une niche écologique spécifique ce
système va être mis à la disposition du reste du monde par le mécanisme de transfert entre les
bactéries.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%