Polycopié du cours


SOMMAIRE
CHAPITRE I: CYCLE ŒSTRAL ...................................................................................................................... 5
I- CARACTERISTIQUES GENERALES DES CYCLES .............................................................................................. 5
1. Terminologie.............................................................................................................................................. 5
2. Durée des différentes phases ..................................................................................................................... 6
3. Les cycles au cours de la vie ..................................................................................................................... 7
II. EVENENEMENTS CELLULAIRES .................................................................................................................... 11
1. Evénements cellulaires ovariens.............................................................................................................. 13
2. Evolution cyclique des voies génitales..................................................................................................... 16
3. Les variations cycliques de l’épithélium vaginal ..................................................................................... 19
III-EVENEMENTS ENDOCRINIENS ET COMPORTEMENTAUX .............................................................................. 22
1. Evénements endocriniens du cycle œstral ............................................................................................... 22
2. L'Oestrus ................................................................................................................................................. 26
3. Cas particuliers ....................................................................................................................................... 29
CHAPITRE II: ENDOCRINOLOGIE DE LA REPRODUCTION : L’AXE GONADOTROPE............... 35
I. LES HORMONES STEROÏDIENNES .................................................................................................................. 36
1. Définition et classification ....................................................................................................................... 36
2. Les voies de la stéroïdogenèse sexuelle ................................................................................................... 38
3. Compartimentation de la stéroïdogenèse sexuelle .................................................................................. 41
4. Rôle physiologique des stéroïdes sexuels ................................................................................................ 44
II- LES GONADOTROPINES HYPOPHYSAIRES ................................................................................................... 50
1. L’hypophyse : anatomie et sécrétions hypophysaires .............................................................................. 50
2. Relation structure-activité des gonadotropines ....................................................................................... 53
3. Rôle physiologique et sécrétion pulsatile des gonadotropines hypophysaires ........................................ 56
La sécrétion pulsatile des hormones gonadotropes hypophysaires ............................................................. 60
III. LES GONADOLIBERINES .............................................................................................................................. 61
1. L’hypothalamus: données anatomiques et hormones hypothalamiques .................................................. 61
2. Rôle physiologique et sécrétion pulsatile du GnRH ................................................................................ 65
3. Régulation de la sécrétion du couple GnRH-LH ..................................................................................... 69
CHAPITRE III: LA FONCTION OVARIENNE ............................................................................................ 77
I. LA GAMETOGENESE FEMELLE ...................................................................................................................... 77
1. Le développement des follicules ovariens ................................................................................................ 77
2. Cinétique de la croissance folliculaire .................................................................................................... 82
3. Régulation du nombre de follicules susceptibles d’ovuler....................................................................... 85
4. L'ovulation ............................................................................................................................................... 95
II. LA FONCTION LUTEALE CYCLIQUE................................................................................................................ 97
1. Caractéristiques morphologiques du corps jaune ................................................................................... 97

3
2. Sécrétion de progestérone et facteurs lutéotropes ................................................................................... 99
III. LES MECANISMES DE LA LUTEOLYSE ....................................................................................................... 100
1. Mise en évidence du rôle de l’utérus dans le processus lutéolytique .................................................... 100
2. Identification des Prostaglandines F2
comme facteur lutéolytique .................................................... 102
3. Mécanismes de régulation de la lutéolyse ............................................................................................. 103
III. LA FONCTION LUTEALE PENDANT LA GESTATION...................................................................................... 105
1. Rôle clé de la progestérone dans le maintien de la gestation ................................................................ 105
2. Mise en évidence du rôle du conceptus dans le maintien de la fonction lutéale .................................... 107
3. Reconnaissance maternelle de la gestation ........................................................................................... 110
CHAPITRE IV. PRODUCTION ET TRANSPORT DES SPERMATOZOÏDES ...................................... 115
I- LA SPERMATOGENESE ................................................................................................................................ 115
1. Anatomie du testicule............................................................................................................................. 116
2. La cytologie et la cinétique de la spermatogenèse ................................................................................ 118
3. Efficacité de la spermatogenèse ............................................................................................................ 126
II. REGULATION DES FONCTIONS TESTICULAIRES ......................................................................................... 127
1. Régulation intra-gonadique des fonctions testiculaires ........................................................................ 127
2. Contrôle endocrinien des fonctions testiculaires ................................................................................... 130
II. TRANSPORT DES SPERMATOZOIDES .............................................................................................. 131
1. Transport épidydymaire ........................................................................................................................ 131
CHAPITRE V. LE COMPORTEMENT SEXUEL ....................................................................................... 133
I. ETHOLOGIE DU COMPORTEMENT SEXUEL .................................................................................................. 133
II. MECANISMES NEUROBIOLOGIQUES DU COMPORTEMENT SEXUEL ............................................................ 136
III. PHYSIOLOGIE DE L’ERECTION ET DE L’EJACULATION ............................................................................... 140
1. Physiologie de l’érection ....................................................................................................................... 140
3. L’éjaculation.......................................................................................................................................... 150
3. Modalités de l’accouplement ............................................................................................................... 153
CHAPITRE VI. RAPPROCHEMENT DES GAMETES ET FECONDATION ........................................ 155
I- ASPECTS ANATOMIQUES ............................................................................................................................. 155
1. Le col de l’utérus ou cervix ................................................................................................................... 155
2. L’utérus ................................................................................................................................................. 156
3. L a trompe utérine ou oviducte .............................................................................................................. 157
II- TRANSPORT ET SURVIE DES SPERMATOZOÏDES DANS LE TRACTUS GENITAL FEMELLE ........................... 157
1. Motricité et transport des spermatozoïdes dans l’utérus ....................................................................... 157
2. Sécrétions utérines lors de la remontée des spermatozoïdes ................................................................. 160
3. Rôle de l’oviducte dans la remontée des spermatozoïdes ...................................................................... 161
III- LA FECONDATION ...................................................................................................................................... 161
1. Pénétration du cumulus oophorus ......................................................................................................... 162
2. Interactions du spermatozoïde et de la zone pellucide .......................................................................... 163
Formation, développement et migration des pronoyaux ........................................................................... 165
IV- TRANSPORT DES ŒUFS ............................................................................................................................ 166

4
1. Motricité tubaire et migration des œufs ................................................................................................. 166
2. Sécrétions tubaires et développement de l’œuf fécondé ........................................................................ 167
3. Motricité utérine et positionnement des œufs ........................................................................................ 167
CHAPITRE VII. DEVELOPPEMENT, IMPLANTATION DU BLASTOCYSTE ET PHYSIOLOGIE
PLACENTAIRE ............................................................................................................................................... 169
I. ASPECTS ANATOMIQUES ET CELLULAIRES DE L’IMPLANTATION ............................................................. 169
1. Développement pré-implantatoire ..................................................................................................... 169
2. Phases et mécanismes cellulaires de l’implantation ............................................................................. 174
II- CONTROLE HORMONAL DE L’IMPLANTATION ............................................................................................. 177
1. Contrôle stéroïdien ............................................................................................................................ 177
2. Réceptivité utérine et activation du blastocyste ..................................................................................... 179
4. Réaction déciduale ................................................................................................................................ 180
III. PHYSIOLOGIE PLACENTAIRE ........................................................................................................... 182
1. Les différents types de placenta ......................................................................................................... 182
2- Fonctions d’échanges du placenta ........................................................................................................ 185
3- Fonction endocrine du placenta ............................................................................................................ 188
CHAPITRE VIII. LE DECLENCHEMENT DE LA PARTURITION ....................................................... 198
I. THEORIE DU BLOCAGE PROGESTERONIQUE ............................................................................................... 198
1. Bases physiologiques ............................................................................................................................. 198
2. Limites de la théorie .............................................................................................................................. 201
II. MISE EN EVIDENCE DU ROLE DU FŒTUS .................................................................................................... 202
1. Observations épidémiologiques ............................................................................................................. 202
2. Preuves expérimentales ......................................................................................................................... 203
3. Sécrétion de cortisol par le fœtus .......................................................................................................... 205
III. MODE D’ACTION DU CORTISOL FŒTAL ...................................................................................................... 207
1. Contrôle de la stéroïdogenèse placentaire ............................................................................................ 207
2. Influence des stéroïdes sexuels sur l’activité contractile du myomètre ................................................. 210
CHAPITRE IX: PHYSIOLOGIE DE LA LACTATION ............................................................................. 221
I. LA MAMMOGENESE ...................................................................................................................................... 221
1. La glande mammaire ............................................................................................................................. 221
2. Les étapes du développement de la glande mammaire .......................................................................... 223
3. Contrôle hormonal de la mammogenèse ............................................................................................... 225
II. LA LACTOGENESE ...................................................................................................................................... 226
1. Les hormones lactogènes ....................................................................................................................... 226
2. Mécanisme d’action des hormones lactogènes ...................................................................................... 227
III. LA LACTATION ........................................................................................................................................... 228
1. La composition du lait et du colostrum ................................................................................................ 228
2. Mécanismes de sécrétion ....................................................................................................................... 231
3. Contrôle hormonal de la lactation ........................................................................................................ 232
4. L’anœstrus de lactation ......................................................................................................................... 235

5
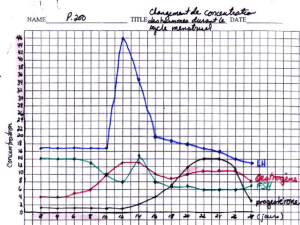
CHAPITRE I: CYCLE ŒSTRAL
Chez la plupart des mammifères, l’ovulation est spontanée: elle a lieu en l’absence
de mâle à des intervalles de temps réguliers caractéristiques de l’espèce.
Deux types de cycles sont distingués: le cycle œstral et le cycle menstruel. Le
cycle œstral est caractérisé par l’apparition périodique d’un comportement
d’œstrus ou d’acceptation du mâle pendant la période qui précède l’ovulation. Au
cours du cycle menstruel, l’activité cyclique des ovaires se manifeste par l’apparition
périodique d’un saignement utérin ou menstruation. L’œstrus et la menstruation
caractérisent respectivement le début du cycle œstral et le début du cycle menstruel.
L’ovulation a lieu au début du cycle œstral et au milieu du cycle menstruel.
Chez certains mammifères appartenant à des ordres divers (lagomorphes,
carnivores), l’ovulation est provoquée par les stimuli tactiles de l’accouplement. Chez
cette espèce, seules les variations des concentrations plasmatiques en stéroïdes
ovariens traduisent les variations cycliques de l’activité ovarienne et permettent
d’identifier les cycles de croissance folliculaire.
I- CARACTERISTIQUES GENERALES DES CYCLES
1. Terminologie
Heape (1900) est le premier à avoir utilisé le terme « œstrus » (adaptation latine du
mot grec oistros) pour désigner la période d’acceptation du mâle. Heape a décrit les
différentes phases du cycle pendant la période d’activité sexuelle en utilisant le
suffixe œstrus et les préfixes pro- met- et di-.
Le proœstrus ainsi défini est la période qui précède l’œstrus et qui correspond à la
croissance folliculaire terminale, l’œstrus étant la période d’acceptation du mâle, du
chevauchement et celle de l’ovulation. C’est au cours du metœstrus que se forment
les corps jaunes à partir des follicules qui ont ovulé. Le diœstrus est caractérisé par
la présence d’un ou plusieurs corps jaunes. En l’absence de fécondation, le corps
jaune régresse, les animaux retournent en proœstrus et ainsi débute un nouveau
cycle.
Une autre terminologie est utilisée pour caractériser les différentes phases du cycle.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
1
/
235
100%









{kind=link}
{kind=link}
{kind=link}