Les mécanismes de l`évolution - Plateforme pédagogique du

3
DOSSIER
Les mécanismes de l’évolution
Ces trois individus appartiennent à un groupe d’individus se
reproduisant entre eux (l’espèce Artedidraco shackletoni, poissons
téléostéens endémiques de l’océan Austral). Ils ont été ramenés par
la mission CEAMARC durant l’hiver 2007-2008, de l’océan Austral,
au large de la Terre Adélie (http://mersaustrales.mnhn.fr/). Les deux
individus du bas ont été capturés au cours d’un même trait de chalut.
Ils présentent entre eux des différences morphologiques nettes, mais
peu de différences moléculaires sur la majeure partie de leurs
génomes. La variabilité qui existe dans les populations et les espèces
DC
Professeur
LES MÉCANISMES
DE L’ÉVOLUTION
Les mécanism es
de l’évolution
La théorie de l’évolution permet
d’articuler les différents pans de
la biologie, ainsi que de comprendre
l’apparition des traits des êtres
vivants et de leur diversité.
La variabilité entre les individus,
la dérive et la sélection sont à
l’origine de la complexité du vivant.
Agnès Dettaï
Maître de conférences au Muséum
national d’Histoire naturelle,
département Systématique
et Évolution, spécialiste de
phylogénie des poissons téléostéens
Variabilité entre individus dans l'espèce Artedidraco shackletoni (© S. Iglesias/CEAMARC/MNHN)
1-LivreDSn°12-bat_HD-1-29:Mise en page 1 10/09/10 10:38 Page14

entre les individus sert de matériau de base à la sélection
naturelle et à la dérive génétique*. Cette variabilité est
acquise de manière majoritairement aléatoire (mutations
et autres modifications du génome) et préexiste à la
sélection et à la dérive. La variabilité est présente à
tous les niveaux dans les organismes, depuis les
molécules jusqu’à la partie du phénotype* visible à l’œil
nu, en passant par la variabilité entre les cellules d’un
même organisme n
DC
Élève
Ces poissons appartiennent à
la même espèce, mais présentent
des différences morphologiques.
Cette variabilité provient surtout
des modifications qui ont eu
lieu, de manière aléatoire,
sur leur patrimoine héréditaire
(gènes et autres séquences) n
15
Les mécanism es
de l’évolution
Variabilité entre individus dans l'espèce Artedidraco shackletoni (© S. Iglesias/CEAMARC/MNHN)
1-LivreDSn°12-bat_HD-1-29:Mise en page 1 10/09/10 10:38 Page15

évolution des organismes à
reproduction sexuée se fonde sur
deux éléments préexistants, l’héri -
tabilité des traits et la variabilité entre
les individus se reproduisant entre eux.
Cette variabilité est mise en place par des
mécanismes aux résultats en très grande
partie aléatoires. Les changements de la
variabilité interindividuelle préexistante, qui
ont lieu au cours du temps (proportions
dans lesquelles sont présentes dans les
populations les différentes versions des gènes
et séquences : les allèles), que ce soit par
hasard (dérive) ou par sélection, conduisent
à des divergences entre les groupes. Ces
différences s’accumulent jusqu’à empêcher la
reproduction entre les individus de deux
groupes évoluant indépendamment, et
continuent à s’accumuler au cours du temps,
pour produire les larges divergences mor -
pho logiques observées actuellement entre
les groupes plus éloignés. Le texte qui suit se
concentre sur la description des mécanismes
de l’évolution chez la plupart des méta zo -
aires (animaux aux cellules dotées de noyaux,
pour la plupart pluricellulaires).
L'HÉRITABILITÉ DES TRAITS
Le concept d’une transmission physique des
caractéristiques des parents lors de la
reproduction est très ancien. Malgré l’ab -
sence de connaissances sur le médium de
cette transmission, ainsi que sur la part
respective des deux parents, la sélection
artificielle a été employée avec succès depuis
plusieurs millénaires par les éleveurs et les
agriculteurs. Les progrès de la compré -
hension des mécanismes de l’hérédité sont
plus récents : seconde moitié du XIXesiècle
(lois de Mendel sur l’hérédité, séparation
des lignées cellulaires somatiques* et ger -
minales*), et surtout recherches au cours
du XXesiècle (l’ADN en tant que support de
l’hérédité, sa structure, les caractéristiques
de sa réplication).
LA VARIABILITÉ ENTRE LES INDIVIDUS
Le concept de transformation au cours du
temps des êtres vivants remonte à bien avant
le XIXesiècle. Cependant, dans les propositions
(qu’elles soient fixistes ou transformistes) qui
précèdent la publication de Darwin, les
variations entre les individus restent
considérées comme des déviations d’un idéal.
Il y a deux nouveautés radicales dans la pro -
position de Darwin par rapport aux
hypothèses précédentes : la place centrale
accordée à la variabilité entre les individus,
qui n’est plus considérée comme un détail
sans importance mais comme le matériau
permettant la transformation des espèces, et
l’idée que la source de la variation est liée au
hasard. La théorie ne se fondait à l’époque
que sur des connaissances fausses ou très
incomplètes des mécanismes sous-jacents.
Nous commençons maintenant à avoir une
bonne idée des mécanismes situés au niveau
moléculaire (génotype*) qui sous-tendent
l’apparition de la variabilité.
La réplication de l’ADN est un processus
semi-conservatif qui permet une copie avec
une très bonne fidélité des brins d’ADN
parentaux. Malgré des processus cellulaires
de correction très performants, la taille même
des génomes fait qu’un très faible taux
d’erreur représentera un nombre conséquent
de différences (les mutations). L’ADN peut
également être endommagé ou remanié de
manière plus ou moins importante à la suite
de processus endogènes (qui ont une cause
interne) ou exogènes (qui ont une cause
externe). D’autres phénomènes peuvent
entraîner, en un seul événement, la modi -
fication d’un grand nombre de paires de
bases, voire de chromosomes entiers : les
insertions et recombinaisons dues à des
éléments transposables* (transposons, rétro -
posons) et à des rétrovirus (qui peuvent être
responsables de certains cancers), les crossing-
over (recombinaisons entre les chromosomes
lors de la méiose), les recombinaisons de
séquences répétées, qui, tous, créent des
allèles ou des combinaisons inédites d’allèles.
Si on considère l’ensemble des modifications
possibles, il y a de nom breuses différences
entre les génomes des parents et ceux des
descendants.
3
DOSSIER
Les mécanismes de l’évolution
Sélection et dérive (© A. Dettaï/MNHN)
L’
1-LivreDSn°12-bat_HD-1-29:Mise en page 1 10/09/10 10:38 Page16

17
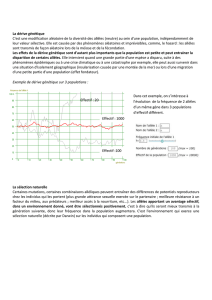
Exemple d’évolution de la proportion de deux allèles
dans une population, pour une petite population
diploïde – ayant un assortiment double de chro -
mosomes – de taille constante (6 naissances,
4 adultes à la reproduction). L’allèle rouge est dominant, et le phénotype
procure aux individus un avantage pour la survie par rapport au
phénotype . Les gamètes sont en grand nombre lors de la reproduction
et dans les mêmes proportions que dans les reproducteurs de la
génération parentale (1/8 pour la génération 1). Par contre, la fréquence
des deux allèles peut changer à cause de biais aléatoires d’échan -
tillonnage, dans les petites populations, dans la transmission des allèles
d’une génération à l’autre (dérive génétique). Il faut noter que même
si le phénotype procure un avantage par rapport au phénotype , il
peut arriver que des individus qui en sont porteurs soient éliminés
(accident, rencontre d’un prédateur, etc.). Inversement, un phénotype
légèrement désavantageux peut persister longtemps n
DC
Professeur DC
Élève
Sélection et dérive (© A. Dettaï/MNHN)
La survie et la
reproduction des
individus dépendent
à la fois des avantages
ou désavantages
que présentent les
différentes versions
des gènes et séquences
(allèles) et des varia-
tions aléatoires des
proportions d’allèles
au cours du temps n
1-LivreDSn°12-bat_HD-1-29:Mise en page 1 10/09/10 10:38 Page17

Le grand nombre de mutations et autres
événements modifiant aléatoirement les
séquences peut sembler présenter un danger
constant et important pour les êtres vivants :
la suppression ou la modification de la
fonction de la protéine résultante sont
nette ment plus probables que son amélio -
ration. Mais seuls les changements dans les
cellules de la lignée germinale peuvent être
transmis à la descendance, et de très
nombreuses modi fications n’ont pas d’impact
sur le phénotype, même si elles peuvent
s’ajouter à la variabilité génétique observée.
• Plus de 98 % du génome humain ne sont pas
codants et plus de 90 % n’ont pas de fonction
connue.
• Le code génétique est redondant : certains
acides aminés peuvent être codés par plusieurs
codons différents. Une mutation qui ne change
pas l’acide aminé codé (mutation synonyme) ne
provoque pas de modification dans la protéine.
• Une mutation qui modifie la séquence pro -
téique ne modifie pas nécessairement le fonc-
tionnement de la protéine.
• Pour de nombreuses protéines, il suffit d’une
seule copie du gène pour que la fonction soit
tout de même assurée, partiellement ou dans
son entier. De plus, certains gènes sont présents
en multiples copies.
• De nombreuses voies métaboliques
com pren nent des redondances. Aussi, une
inac tivation complète de la protéine codée
par un gène peut ne pas avoir d’impact sur la
viabilité de l’organisme touché.
Certaines modifications du matériel génétique
ont une influence sur le phénotype, et seront
soumises à sélection. Les changements se
produisent sur de très grandes durées, de très
grands nombres de groupes et d’individus, et
les génomes eux-mêmes sont de grande taille.
De très nombreuses altérations, duplications,
mutations se sont donc généralement produites
pour la plupart des différences que nous
observons.
• La redondance (copies multiples, voies
multiples, hétérozygotie*) permet la mo di -
fication d’une des copies sans que cela soit
préjudiciable à l’organisme. Certaines copies
acquièrent ainsi au fil du temps de nouvelles
propriétés, voire une nouvelle fonction dans
l’organisme.
• Chez les eucaryotes*, la modularité de la
plupart des gènes (structure en introns et exons,
c’est-à-dire alternance dans un gène de régions
codantes et non codantes) permet d’obtenir
plusieurs protéines à partir d’un seul gène, en
combinant des exons différents, mais aussi de
créer de nouveaux gènes, à partir d’une recom -
binaison d’exons de gènes existants.
• Les séquences régulatrices ne sont pas
codantes mais elles déterminent dans quel type
de cellules, quelle partie du corps et à quel
stade du développement les gènes sont
exprimés. Ce type de modifications est proba -
blement l’une des clés pour comprendre la
diversité de forme des organismes, car leur
modification peut avoir des conséquences
importantes sur l’organisme.
3
DOSSIER
Les mécanismes de l’évolution
zoom SUR
Plusieurs points méritent d’être rappelés à propos de la sélection, une des parties les plus
emblématiques de la théorie de l’évolution. La sélection assure un tri sélectif des individus en
fonction de leur phénotype. Celui-ci comprend les caractéristiques morphologiques et
comportementales, mais également l’ensemble des molécules qui assurent le fonctionnement
(protéines...). Il n’est possible de dire qu’un individu est plus adapté qu’un autre (avantage
sélectif) que pour un environnement donné. Si l’environnement (milieu, compétition avec des
individus de la même espèce ou avec d’autres espèces) change, les valeurs sélectives changent. Plus
la différence entre la valeur sélective des deux allèles dont les porteurs sont en compétition est
grande, plus l’effet (la sélection) sera fort n
La sélection
Spéciation allopatrique (© A. Dettaï/MNHN)
1-LivreDSn°12-bat_HD-1-29:Mise en page 1 10/09/10 10:38 Page18
 6
7
8
6
7
8
1
/
8
100%