1. Hormones effectrices

Physiologie endocrinienne
PHYSIOLOGIE DU SYSTEME ENDOCRINIEN – GENERALITE
I. Définition
1. Système endocrinien
- Il assure essentielle de communication comme le système nerveux mais sur d’autres bases
et différent en termes de délégation.
- Si le système nerveux communique c’est dans le but de coordonner la physiologie de
l’organisme. Le système endocrinien lui assure la communication et le développement
de l’organisme.
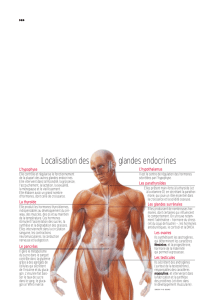
- Il est constitué de glandes endocrines plus ou moins anatomiquement isolés les unes des
autres :
o Au niveau de l’encéphale on retrouve l’hypothalamus et l’hypophyse (système
hypotalamo-hypophysaire).
o Au niveau du cou, la thyroïde et les parathyroïdes.
o Les glandes surrénales coiffent les reins.
o Le pancréas.
o Les gonades (ovaires ou testicules).
- A côté de ces glandes on retrouve des cellules endocrines disséminée:
o Au niveau du tube digestif.
o Au niveau du système rénal.
2. Hormones
- Ces glandes et ces cellules endocrines sécrètent des molécules de signalisation (comme
dans le système nerveux) correspondant à des hormones (et non des neurotransmetteurs)
qui agissent à distance parce qu’elles sont transportées par le sang vers les tissus cibles.
- On peut étendre cette notion d’hormone en incluant les molécules qui agissent par voie
paracrine qui ne sont pas transportées par le sang mais qui agissent sur les cellules
voisines de la cellule sécrétrice. On parle alors de para-hormone.
- A l’extrême il existe des cellules sécrétrices qui agissent sur elle-même, par voie
autocrine. On parle alors d’auto-hormone.
- Dans le cas particulier où l’hormone est sécrétée par un neurone. On parle alors de
neurohormone.
- Ces différentes types d’hormones agissent sur des tissus cible via des récepteurs
spécifique de l’hormone. Cette liaison se traduit par des effets biologiques consécutifs à
des modifications l’activité cellulaire de ces tissus.
- Remarques :
o Certaines hormones sont aussi des neurotransmetteurs : exemple de la
noradrénaline.
o Une glande peut sécréter plusieurs hormones.
o Une même hormone peut être produite par diverses glandes.
o Une hormone peut agir sur des tissus cibles différents et donc intervenir dans
diverses fonctions.
II. Nomenclature – hiérarchie
1. Hormones effectrices
- On parle d’hormones effectrices quand elles sont à l’origine directe d’effets biologiques
en agissant sur les tissus cibles.
- Ces hormones portent un nom proche de la glande qui les sécrète :
o La thyroïde sécrète les hormones thyroïdiennes.
o Les corticosurrénales sécrètent les hormones cortico-surrénaliennes.

2. Les hormones de rang supérieur
- Ces hormones là n’agissent pas sur des tissus cibles. Elles sont des actions indirectes sur
les tissus cibles par l’intermédiaire d’autres hormones.
- Il y en a deux types :
o Les hormones glandulotropes qui agissent sur des glandes endocrines lesquels
libèrent les hormones effectrices.
Exemple : hormones de l’hypothalamus.
Pour les nommer on utilise le suffixe « tropine ». Exemple : l’axe thyrotropine et
l’axe corticotropine.
o Les facteurs de stimulation ou d’inhibition des hormones glandulotropes.
S’il s’agit de facteurs de stimulation on utilise les suffixes « stimuline » ou
« libérine ». Exemple : dans l’axe corticosurrénalien il y a une corticostimuline.
S’il s’agit de facteurs d’inhibition on utilise le suffixe « statine ». Exemple : la
somatostatine.
- Ces hormones de rang supérieur, ce sont elle qui en plus de l’effet sur la glande, assure la
trophicité et la croissance des glandes endocrines (qui sont en quelques sortes leur tissu
cible).
- On peut avoir en pathologie :
o Des hypertrophies des glandes endocrines lorsqu’il y a une hypersécrétion des
facteurs de stimulation.
o Des hypotrophies en cas d’hypersécrétion de facteurs d’inhibition.
III. Structure des hormones
- Si on regarde leur structure on peut distinguer trois groupes :
o Les hormones aminées.
o Les hormones stéroïdes.
o Les hormones peptidiques.
1. Les hormones aminées
- Les hormones aminées sont toutes dérivées d’un acide aminé appelé tyrosine.
- C’est le cas des hormones thyroïdiennes : la tri-iodo-thyronine (T3) et la tétra-iodo-
thyronine (T4 ou thyroxine).
- Les catécholamines (dopamine, noradrénaline et adrénaline) peuvent avoir une fonction
d’hormone :
o Noradrénaline et adrénaline sont des hormones de la médullo-surrénale.
o La dopamine est un facteur inhibiteur d’origine hypothalamique de sécrétion de la
prolactine. C’est la prolactine-statine.
2. Les hormones stéroïdiennes
- Les hormones stéroïdiennes ont comme précurseur de synthèse le cholestérol.
- Ce sont les hormones des corticostéroïdes sécrétés par les corticosurrénales. Ils sont de
trois ordres selon la partie des corticosurrénales :

o L’aldostérone est une hormone minéralo-corticoïde qui intervient sur l’équilibre
hydro-électrolytique.
o La cortisone et la cortico-stérol qui eux sont des glucocorticoïdes. Ils interviennent
dans le métabolisme énergétique.
o Les androgènes-surrénaliens.
- La testostérone sécrétée par le testicule.
- Les œstrogènes et la progestérone produite par les ovaires.
- La vitamine D peut également être considérée comme une hormone. Elle intervient dans
le métabolisme phosphocalcique.
3. Les hormones peptidiques
- Toutes les hormones non aminées et non stéroïdiennes ont une structure de peptide.
- Il peut également s’agir de gluco-protéine : structure mixte.
- Elles sont souvent sécrétées sous forme de précurseurs qui sont des chaines d’acides
aminés contenant l’hormone qui seront clivés par la suite. On parle alors de pro-
hormones.
- Il peut aussi y avoir des précurseurs de précurseurs appelé pré-pro-hormone.
- Exemple de l’insuline (pré-pro-insuline et pro-insuline).
IV. Les hormones dans le sang
1. Transport
- Le transport des hormones dépend si l’hormone est :
o Hydrophobe.
o Hydrophile.
- Plus elle est hydrophile plus elle est susceptible de circuler sous forme libre.
Schématiquement les hormones peptidiques sont souvent des domaines hydrophiles et
circulent donc libres.
- Lorsqu’elle est hydrophobe elle sera transportée par des protéines de transport.
Par contre les hormones aminées ou stéroïdes sont plutôt hydrophobes et circulent liées à
des protéines de transport telles que l’albumine ou des molécules plus spécifiques
appelées binding-protéine.
Dans ces cas là c’est la forme libre (même si elle est en faible concentration) qui est active
sur les tissus.
Bien entendu au fur et à mesure qu’un certain nombre de molécule libre ont été utilisée,
les protéines de transport libère les autres molécules (équilibre).
2. Durée de vie
- La durée de vie va être différente suivant la liposolubilité ou l’hydrosolubilité des
hormones.
- De manière générale les hormones hydrosolubles (peptidiques) peuvent être dégradées
rapidement. Les réponses qu’elles fournissent lorsqu’elles sont sécrétées sont donc de
courtes durées.
- Les hormones hydrophobes (aminées et stéroïdiennes) sont moins facilement dégradable.
Leurs actions sont donc plus longues. Par exemple les hormones stéroïdiennes ne
produisent leur effet que quelques jours après injection.
V. Les modes d’actions des hormones
1. Mode d’action cellulaire
- Ces hormones agissent sur des récepteurs auxquels elles se lient pour donner un système
effecteur qui déclenche les effets biologiques.
- En principe les récepteurs sont spécifiques de l’hormone mais la reconnaissance n’est pas
exclusive pour l’hormone. Les récepteurs pour l’hormone peuvent également lié d’autres
ligands, en particulier les métabolites de l’hormone donnant des effets métaboliques
moindres.

- Certains récepteurs lient d’autres hormones.
- Les récepteurs peuvent lier des molécules pharmacologiques, des antagonistes de
synthèse.
- Il existe deux grands types de récepteurs :
o Des récepteurs intracellulaires.
o Des récepteurs membranaires.
- Les hormones hydrophobes rentrent dans la cellule pour aller agir sur des récepteurs
intracellulaires (leur liposolubilité permet de franchir la membrane cellulaire).
- Les hormones hydrophiles agissent sur des récepteurs membranaires.
- Ces récepteurs intracellulaires peuvent être des protéines du cytosol ou ils peuvent être
des récepteurs intranucléaires. Quand le complexe hormone-récepteur est formé, il se lie à
l’ADN pour induire une synthèse protéique.
- Les récepteurs membranaires se divisent en deux catégories :
o Les récepteurs couplés aux protéines G possédants 7 domaines membranaires.
o Les récepteurs couplés à des enzymes ne possédant qu’un seul domaine
membranaire. C’est le cas de deux hormones importantes :
o L’insuline.
o L’hormone de croissance.
Si l’enzyme correspond à une protéine kinase l’effet passe par la phosphorylation
de substrat.
Cas particulier : l’hormone pénètre dans le tissu cible par endocytose après s’être lié à un
récepteur (endocytose du complexe hormone-récepteur).
2. Effets biologiques
- Les effets biologiques des hormones sont des effets qui permettent le contrôle du
métabolisme par :
o Des actions de régulation de synthèse d’enzyme.
o Des modifications de configuration d’enzyme.
o Des modifications de quantité disponible de substrat.
- Il existe des régulations de synthèse des récepteurs par la concentration d’hormone. Par
exemple quand la concentration d’insuline augmente, le nombre de récepteurs diminue
(autorégulation par feed-back).
- Il y a la possibilité de modulé les récepteurs d’une hormone par une autre hormone. Par
exemple T3 augmente le nombre de récepteurs à l’adrénaline sur le cœur.
VI. La sécrétion hormonale
1. Décours temporel
- L’hormone n’est généralement pas sécrété de manière continue, mais par des bouffées
plus ou moins fréquentes. De ce fait la concentration plasmatique des hormones n’est pas
stable, encore moins contrôlée de manière homéostatiques.
- En plus de ça les bouffées de sécrétions n’ont pas la même amplitude : on peut voir des
pics de sécrétion. Il y a des variations cycliques de ces taux. Exemple de l’hormone de
croissance sécrétée avec un grand pic en début de sommeil.
2. Contrôle
- Il existe différents contrôle de la sécrétion qui s’intègre (soit dans le sens de stimulation,
soit dans le sens d’inhibition) :
o Ceux contrôlés par le système nerveux.
o Ceux contrôlés par d’autres hormones.
- Les sécrétions contrôlées par le système nerveux correspondent par exemple à celle de la
médullosurrénale contrôlée par le système orthosympathique.
- L’hypothalamus intervient également il correspond au « cerveau endocrinien ».
- Les hormones peuvent contrôlées la sécrétion d’autres hormones à l’aide de feedbacks,
de rétrocontrôles, positifs ou négatifs. Le feedback négatif prédomine.

- Les feedback négatifs peuvent venir de l’hormone effectrice pour aller sur les hormones
de rangs supérieurs.
- Si au contraire il n’y a pas assez d’hormone effectrice il peut y avoir :
o Stimulation de sécrétion de stimuline par l’hypothalamus.
o Ou une stimulation de sécrétion de tropine par l’hypophyse.
VII. Physiopathologie
- On peut aboutir à une physiopathologie en cas :
o Diminution ou augmentation de l’action hormonale.
o Altération des boucles de régulation
- La diminution ou l’augmentation de l’action hormonale a lieu lorsque la glande
endocrine n’est pas assez ou trop stimulée.
o Lorsqu’il y a une pathologie au niveau des glandes sécrétrices de l’hormone, trop
nombreux (hyperplasie) ou pas assez nombreuses (hypoplasie) ou pas du tout de
cellules (aplasie).
o Lorsque les glandes sont en position ectopiques (pas à leur place) il y a alors une
sécrétion ectopique.
o Un défaut de réponse des cellules sécrétrices aux stimuli.
o Un problème de synthèse de l’hormone.
o Un problème de transport intracellulaire (de sécrétion).
o Un problème de transport sanguin.
o Un problème au niveau des récepteurs de la cellule cible.
- L’altération des boucles de régulation peut avoir lieu dans le système hypothalamo-
hypophysaire. Par exemple :
o L’augmentation de sécrétion de stimuline (hypothalamus) donne lieu à une
augmentation de sécrétion de tropine (hypophyse) qui donne lieu à une
augmentation de sécrétion d’hormone effectrice (glande endocrine) et donc à une
augmentation des effets (tissu cible).
o En cas d’augmentation des effets, un système de feedback négatif est appliqué sur
la glande endocrine et sur la sécrétion des hormones de rang supérieur.
o Si ces systèmes de régulations, ces feedback, sont défectueux ont aboutit à un
excès des effets biologiques.
LE SYSTEME HYPOTHALAMO-HYPOPHYSAIRE
I. Rappels anatomo-histologiques
Hypothalamus
- C’est une petite structure à la base du cerveau ne pesant que 4 grammes.
- Elle constitue les parois du 3ème ventricule.
- Elle est reliée à sa base à l’hypophyse. La partie qui relie les deux est la tige pituitaire.
- L’éminence médiane correspond à la partie inférieure de l’hypothalamus.
Hypophyse
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
1
/
46
100%