Anatomie comparée du « chiasma optique » chez les Vertébrés

0
UNIVERSITE DE NANTES
FACULTE DE MEDECINE
MASTER I SCIENCES BIOLOGIQUES ET MEDICALES
UNITE D’ENSEIGNEMENT OPTIONNEL
MEMOIRE REALISE dans le cadre du CERTIFICAT d’ANATOMIE,
d’IMAGERIE et de MORPHOGENESE
2004-2005
UNIVERSITE DE NANTES
Anatomie comparée du « chiasma optique » chez les
Vertébrés
Par
Salaud Céline
LABORATOIRE D’ANATOMIE DE LA FACULTE DE MEDECINE DE NANTES
Président du jury : Pr. J. LE BORGNE
Vice-Président : Pr. J.M. ROGEZ
Enseignants :
• Pr. O. ARMSTRONG
• Dr. O. BARON
• Pr. C. BEAUVILLAIN
• Dr F. CAILLON
• Pr. P. COSTIOU
• Pr. D. CROCHET
• Dr J. DELECRIN
• Dr. H. DESAL
• Pr. B. DUPAS
• Dr E. FRAMPAS
• Dr A. HAMEL
• Pr. Y. HELOURY
• Dr M.D. LECLAIR
• Pr. P.A. LEHUR
• Pr. N. PASSUTI
• Pr. R. ROBERT
• Pr. D. RODAT
• Dr VALETTE
Laboratoire : S. LAGIER et Y. BLIN - Collaboration Technique

1
UNIVERSITE DE NANTES
FACULTE DE MEDECINE
MASTER I SCIENCES BIOLOGIQUES ET MEDICALES
UNITE D’ENSEIGNEMENT OPTIONNEL
MEMOIRE REALISE dans le cadre du CERTIFICAT d’ANATOMIE,
d’IMAGERIE et de MORPHOGENESE
2004-2005
UNIVERSITE DE NANTES
Anatomie comparée du « chiasma optique » chez les
Vertébrés
Par
Salaud Céline
LABORATOIRE D’ANATOMIE DE LA FACULTE DE MEDECINE DE NANTES
Président du jury : Pr. J. LE BORGNE
Vice-Président : Pr. J.M. ROGEZ
Enseignants :
• Pr. O. ARMSTRONG
• Dr. O. BARON
• Pr. C. BEAUVILLAIN
• Dr F. CAILLON
• Pr. P. COSTIOU
• Pr. D. CROCHET
• Dr J. DELECRIN
• Dr. H. DESAL
• Pr. B. DUPAS
• Dr E. FRAMPAS
• Dr A. HAMEL
• Pr. Y. HELOURY
• Dr M.D. LECLAIR
• Pr. P.A. LEHUR
• Pr. N. PASSUTI
• Pr. R. ROBERT
• Pr. D. RODAT
• Dr VALETTE
Laboratoire : S. LAGIER et Y. BLIN - Collaboration Technique

2
ANATOMIE COMPAREE DES VOIES OPTIQUES
Je tiens particulièrement à remercier :
Monsieur le Professeur R. ROBERT, pour nous avoir attribuer un sujet de mémoire
qui nous tient à cœur, mais également et surtout pour son soutien et ses conseils précieux,
Monsieur le Professeur G. PRADAL, pour les coupes histologiques et le temps qu’il
nous a accordé, mais aussi pour nous avoir fourni un poulet,
Monsieur le Professeur P. COSTIOU et l’école vétérinaire pour leur disponibilité,
leurs conseils et leur accueil et pour nous avoir fourni les animaux,
Messieurs S. LAGIER et Y. BLIN, personnel du laboratoire d’anatomie, pour nous
avoir suivi tout au long de l’année, pour leur disponibilité et leur soutien,
Madame A. HIVONNAIT, chirurgie expérimentale et animalerie, pour nous avoir
aidé,
Monsieur P. JOLIVEL, informaticien, pour son aide.

3
SOMMAIRE
I - Introduction
II - Rappels embryologiques
III - Elément de classification des vertébrés
1) Classification rudimentaire des Cordés
2) Apparition des Vertébrés au cours des âges géologiques
IV - Rappels anatomiques
1) Poissons
2) Amphibiens et Reptiles
3) Mammifères, Homme
A- Le nerf optique
B- Le chiasma optique
C- Le tractus optique
D- Le corps géniculé latéral
E- Les radiations optiques
F-Le centre cortical visuel
G- Le Chien
4) Oiseaux
V - Matériels et méthodes
1) Pièces anatomiques
2) Instruments
3) Dissections
A- Homme
B- Chien
C- Poule
D- Poisson
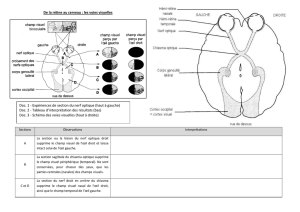
VI - Approche microscopiques du chiasma optique
A- Connexions des rétines avec les centres nerveux chez les Vertébrés
B- Systématisation du chiasma optique chez l’Homme
C- Approche histologique du chiasma optique chez le Poulet
D- Approche histologique du chiasma optique chez le Renard et le Lapin
E- Radioautographie du mésencéphale d’un Amphibien
VII - Discussion
A- La présence du « chiasma », ou entrecroisement chez les Vertébrés
B- Y’ a-t-il un échange fonctionnel des fibres rétiniennes chez tous les
Vertébrés
C- Voies visuelles accessoires
D- Cas extrême du Caméléon
E- Pourquoi un chiasma ?
F- L’œil et la vision chez les Mammifères
VIII – Conclusion
IX – Bibliographie

4
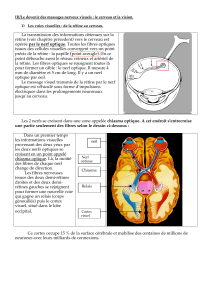
I - Introduction
Vue caudale d’un encéphale d’Homme, voies optiques
La dénomination de chiasma optique provient directement de sa forme. En effet, le chiasma
est comparé à la lettre grecque khi correspondant au X de l’alphabet grec et représente
l’entrecroisement « morphologique » des deux nerfs optiques.
Cependant, la réelle signification du chiasma n’est elle pas un échange fonctionnel des fibres
rétiniennes au sein de ce dernier ? Il ne viendrait pas à l’esprit de considérer comme chiasma
le croisement de l’artère utérine avec l’uretère et pourtant leur croisement est réel.
Cet échange, bien connu chez l’Homme, est il retrouvé chez les autres Vertébrés ?
Nous allons donc tenter de démontrer si l’on retrouve un chiasma morphologique ou un réel
échange fonctionnel des fibres et de savoir si cela est corrélé au mode de fovéation des
différents Vertébrés.
Notre étude comparative portera donc sur l’Homme, le Chien, le Poisson, la Poule et quelques
planches sur les Amphibiens et les Reptiles.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
1
/
40
100%