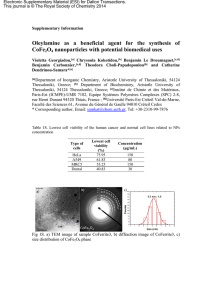
senescence

!ynamics and plas"ci# of $lomeres :
%onsequences in cancer and aging &
'(ronique Gire&
)RBM-CNRS, Montpe*ier&
+er[email protected].,&

age !
incidence!
mice!
human!
)ancer incidence rise exponen"a*y wi- age&
0 1.5 3 ! ! 60 ! !120!
Age is the largest simple risk factor !

)e*ular .ansforma"on is a mul"-s$p process&
ADN!
ADN !
«"tumoral"»!
ADN!
viral infection!
UV radiation!
inherited genetic
mutations!
chemical carcinogens!
DNA Repair!
apoptosis!
I
N
I
T
I
A
T
I
O
N
T
R
A
N
S
F
O
R
M
A
T
I
O
N
oncogenes!
chronic inflammation!
tumor suppressor !
genes!
telomerase !
mutations!
mutations!

/a*marks of cancer&
comprise eight biological capabilities acquired during the multistep development of human tumors !
Hanahan& Weinberg, Cell 2000, 2011!

Organisms wi- renewable "ssues had 0 &
evolve mechanisms 0 prevent cancer&
One such mechanism is ce*ular senescence,&
1hich irreversibly arrest -e grow- of ce*s &
at risk of neoplas"c .ansforma"on&
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
1
/
62
100%