sur une nouvelle espèce de porphyra (p. drach/1 j

Vie Milieu, 1978-79, Vol. XXVIII-XXIX, fasc. 3,
sér.
AB,
pp.
425-435.
SUR
UNE
NOUVELLE
ESPÈCE
DE
PORPHYRA
(P.
DRACH/1
J.
FELDM)
ET
REMARQUES
SUR
L'ÉVOLUTION
DES
PLASTES
CHEZ
LES
RHODOPHYCÉES
(BANGIOPHYCIDÉES
ET
ACROCHAETIALES)
par
Jean
FELDMANN
t
Laboratoire
de Biologie végétale
marine,
Université
Pierre
et Marie Curie,
7,
Quai
Saint-Bernard,
75230 Paris
Cedex
05
ABSTRACT
Description
of
a
new
species
of
Porphyra
growing
on
Laminaria
hyperborea
in
the
Channel.
It
is
distinguished
by
its
very
thin
thallus
and
particularly
by
its
parietal
chromatophore
with
a
pyrenoid.
Only
monospores
(neutra!
spores)
have
been
observed.
Morphological
evolution
of
chromatophores
among
the
Bangiophy-
cideae
and
the
more
primitive
Florideophycideae
(Acrochaetiales
and
Nemalionales)
can
be
traced
froqt
cells
with
only
one
central
(focal)
chromatophore
with
a
pyrenoid
to
cells
with
severa!
chromatophores
without
pyrenoid
in
each
cell,
as
it
is
the
case
among
the
more
advanced
Florideophycideae.
Sorne
examples
of
intermediate
forms
are
described.
Latin
diagnosis
of
the
previously
described
genus
Rhodothamniella
characterized
by
its
3-12
parietal
chromatophores
with
a
pyrenoid
in
each
cell,
is
given.

426
J.
FELDMANN
INTRODUCTION
En
août
1948, le
professeur
Pierre
DRACH
récoltait,
au
cours
d'une
plongée
en
scaphandre
autonome
dans
la
baie
de
Morlaix,
devant
Primel,
par
une
dizaine
de
mètres
de
fond,
un
échantillon
de
Laminaria
hyperborea
(Gunner)
Foslie
portant,
sur
la
marge
de
sa
lame,
des
thalles
d'un
petit
Porphyra
qui,
par
ses
caractères,
me
parut
distinct
des
autres
espèces
de
ce
genre.
Retrouvé
ensuite,
en
septembre
à
très
basse
mer
et
en
plongée
dans
les
parages
de
Duon
et
au
Beclem
(Baie
de
Morlaix),
je
signalai
cette
espèce
dans
l'Inventaire
de
la
Flore
marine
de
Roscoff
(1954)
sous
le
nom
de
Porphyra
drachii nov.
sp.
sans
toutefois
l'accompagner
d'une
diagnose.
Tous
les
échantillons
récoltés
étant
stériles
ou
producteurs
seulement
de
monospores,
j'espérais
trouver
ultérieu-
rement
des
plantes
sexuées
qui
m'auraient
permis
de
mieux
pré-
ciser
ses
affinités.
Ce
ne
fut
pas
le
cas.
La
publication
de
ce
fascicule
de
Vie
et
Milieu
dédié
au
Profes-
seur
P.
DRA
CH
me
donne
l'occasion
de
valider
cette
espèce
nouvelle
qui,
bien
que
ses
organes
sexués
soient
inconnus,
me
paraît
bien
caractérisée.
DESCRIPTION
DU
PORPHYRA
DRACHII
NOV.
SP.
DIAGNOSE.
Thallus
monostromaticus,
usque
ad
45
mm
altus, tenuis,
22-28
J,J.m
crassus, oblongus, infra cordiformis,
margine
integra,
vix
undulata, cellulis
subquadratis
12-17
JJ.m
latae plus
minusve
regulariter dispositis.
Chromatophorum
in
quaque
cellula
unicum,
in lamina parie-
tale stellatim lacerata et pyrenoide conspicuo
productum.
Monosporae e
transformatione
cellularum
marginis
ortae.
Propagatio sexualis ignota.
Habitat in oris Armaricae
ad
frondes
Lamanariae
et
Saccorhizae.
Typus
in
Herb. -J.
FELDMANN
(PC.)
Le
Porphyra
drachii (Fig.
1)
forme
des
thalles
d'un
rose
pâle,
oblongs
et
à
marge
entière
et
plus
ou
moins
ondulée,
à
peine
stipités
et
à
base
cordiforme,
atteignant
45
mm
de
haut
et
27
mm
de
large.
Ce
thalle
est
monostromatique,
plus
mince
que
celui

PORPHYRA
DRACHII,
NOV. SP. 427
des
autres
Porphyra
de
nos
côtes
et
fixé
au
substrat
par
les
rhi-
zoïdes
issus
des
cellules
inférieures
du
thalle.
Dans
ses
parties
moyennes,
il
mesure
22-28 !-lill
d'épaisseur.
Les
cellules
vues
de
face,
disposées
plus
ou
moins
régulièrement,
sont
polygonales,
rectangulaires
ou
arrondies
et
mesurent
12-17 !-lill
de
diamètre.
En
coupe,
elles
apparaissent
situées
dans
une
paroi
générale
....._.
lem
FIG.
1. -
Porphyra
drachii
J.
Feldm.
relativement
épaisse
et
sont
sensiblement
aussi
hautes
que
larges
(Fig. 2
A-E).
Chaque
cellule
renferme
un
plaste
pariétal
en
forme
de
lame
découpée
en
étoile
dont
les
branches
s'appliquent
contre
la
paroi
cellulaire
dans
le
cytoplasme
pariétal.
Ce
plaste
est
pourvu
d'un
pyrénoïde
central
bien
visible.
Le
noyau,
difficilement
visible
sur
le
vivant,
est
situé
contre
la
face
interne
du
plaste
pariétal
au
voisinage
du
pyrénoïde.
Les
seuls
organes
reproducteurs
observés
sont
des
monospores
(spores
neutres
ou
gonidies).
Les
monosporocystes
résultent
de
la
transformation
directe
des
cellules
végétatives
de
la
marge
de
la
fronde
qui
deviennent
plus
volumineuses,
s'arrondissent
et
se
chargent
de
petits
grains
d'amidon
floridéen.
Les
monospores
sont
mises
en
liberté
par
gélification
des
parois
du
thalle;
elles
présen-
tent
alors
pendant
quelques
temps
des
mouvements
amiboïdes
(Fig. 3,
A,
B)
avant
de
se
développer,
souvent
in situ.
Il
y a
d'abord
allongement
de
la
spore
qui
s'est
entourée
d'une
paroi
mince,
apparition
d'un
rhizoïde,
tandis
que
la
spore
se
cloisonne
perpendiculairement
à
l'axe
déterminé
par
l'orientation
du
rhizoïde
(Fig. 3,
C,
D)
pour
former
un
filament
dressé.
Par
de
nouveaux
cloisonnements
perpendicul~ires
aux
précédents,
ce
filament
de-
viendra
un
nouveau
thalle
en
lame.
Comme
on
l'a
vu
plus
haut,
je
n'ai
pas
observé
d'organes
reproducteurs
sexués.
Le
Porphyra
drachii se
distingue
des
autres
espèces
de
nos
côtes
par
son
thalle
très
mince,
ses
cellules
subcubiques
et
non

428
J.
FELDMANN
B
L
20
l'
~L
c
FIG.
2. -A-E.
Porphyra
drachii;
A,
B :
coupes
transversales
de
la
fronde;
C :
cellules
vues
de
face;
D :
cellule
vue
en
coupe;
E :
cellule
du
bord
du
thalle;
F :
cellule
de
Porphyra
leucosticta
vue
de
face.

PORPHYRA
DRACHII,
NOV.
SP.
429
A
B
FIG.
3. -
Porphyra
drachii.
A
et
B :
monospores
libérées
présentant
des
mou-
vements
amiboïdes;
C
et
D :
monospores,
entourées
de
la
paroi
gélifiée
du
sporocyste,
se
développant
in
situ.
allongées
perpendiculairement
au
plan
du
thalle.
Il
n'est
pas
sans
rappeler
un
peu
le
Porphyra
leucosticta
qui
vit
à
très
basse
mer
mais
il
s'en
distingue
nettement
par
son
plaste
en
lame
pariétale
découpée,
appliqué
contre
la
paroi
de
la
cellule
dans
le
cytoplasme
pariétal.
Tous
les
autres
Porphyra
possèdent
un
plaste
central
(ou
focal) avec
des
branches
rayonnantes
à
partir
du
pyrénoïde,
dans
toutes
les
directions
de
l'espace
(Fig. 2
F).
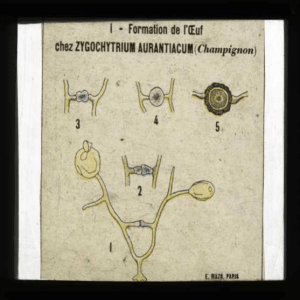
EVOLUTION
MORPHOLOGIQUE
DE
L'APPAREIL
PLASTIDIAL
DES
RHODO-
PHYCÉES.
La
morphologie
de
l'appareil
plastidial
des
Rhodophycées,
très
variable
comme
dans
d'autres
embranchements
d'Algues,
constitue
un
caractère
systématique
important.
Chez les
Bangiophycidées,
comme
chez
les
Florideophycidées,
on
peut
assister
à
une
évolution
progressive
de
l'appareil
plastidial
depuis
la
structure
archéoplas-
tidiée,
considérée
comme
primitive,
jusqu'à
la
structure
néoplasti-
diée.
La
première
est
caractérisée
par
un
plaste
unique
focal
avec
un
pyrénoïde
central
et
des
branches
rayonnantes
dans
toutes
les
directions.
Dans
la
seconde,
chaque
cellule
renferme
de
nombreux
plastes
pariétaux
sans
pyréiioïde.
Le
premier
type
s'observe
notamment
chez
les
Porphyridiales
(Porphyridium),
les
Bangiales
(Porphyra, etc.), les
Acrochaetiales
(Acrochaetium sp.)
et
certaines
Némalionales
(Nemalion, Liagora).
Le
second
type
qui
caractérise
certaines
Bangiophycidées,
Aero-
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%