Olivier Lespinet - Université Paris-Sud

Orsay
N° d’ordre :
UNIVERSITE DE PARIS-SUD
U.F.R. SCIENTIFIQUE D’ORSAY
THESE
Présentée
Pour obtenir
Le GRADE de DOCTEUR EN SCIENCES
DE L’UNIVERSITE PARIS XI ORSAY
par
Olivier Lespinet
Sujet :
La famille des gènes Snail.
Caractérisation de deux nouveaux membres chez le mollusque Patella vulgata.
Hypothèse sur leur fonction ancestrale chez les Bilateria.
Soutenue le lundi 15 janvier 2001, devant la Commission d’examen
M. Jo van den Biggelaar Rapporteur
M. Michel Volovitch Rapporteur
M
me Bernadette Limbourg-Bouchon Examinateur
M
me Angela Nieto Examinateur
M. André Adoutte Examinateur
M. Michel Cassan Examinateur

Il n'y a pas de vie sans risque…
La vie sans risque,
c'est une sorte de mort permanente,
c'est une vie d'esclave.
Léo Lagrange, 1940

Remerciements
J'aimerais tout d'abord remercier les membres du jury pour leur enthousiasme, leur patience et leur
disponibilité.
J'aimerais également remercier ici toutes les personnes qui ont contribué de quelque façon que ce soit
au bon déroulement de ma thèse. Je remercie tout particulièrement André Adoutte et Michel Cassan
pour leurs conseils, leurs encouragements et leur soutien tout au long de ces années.
Je ne voudrais pas non plus oublier ceux qui ont partagé avec moi les moments d'exaltation et parfois
de doute qui se sont succédés durant ces quatre années. Que ce soit à Gif-sur-Yvette: Benjamin, Cyril,
Guillaume, Jean-François, Martine, Michel, Nicolas et Renaud, à Roscoff: Marie ou bien encore à
Utrecht: André, Anke, Eric, Hans, Lex, Pascal et Wim.
Je tiens également à remercier ceux qui au cours de ces années ont toujours essayé de satisfaire ma
curiosité, m'ont fait partager leur passion et m'ont soutenu dans toutes mes initiatives: Jo van den
Biggelaar, Claus Nielsen, Marguerite Picard et Michel Termier.
Je voudrais également remercier ceux qui m'ont accueillis chez eux et m'ont fait la preuve de leur
amitié durant mes années orcéennes et giffoises: Fatima, Marie-Hélène, Véronique, Henri et la joyeuse
troupe de la rue du Général Duchesne et plus récemment, Michel, Marie-Claire, Ambroise et Marie.
Sans oublier Anne, pour notre amitié, nos rêves de révolution, son pastis et sa spécialité au miel et au
melon !
J'aimerai enfin terminer par remercier Micheline, Gilbert et Hélène pour tout ce qu'ils ont su me
donner, ainsi que tous ceux du grand-ouest (Bordelais, Nantais, Niortais, Oulmais, Rethais, etc.) qui
dans les bons comme dans les mauvais coups de vent ont toujours su rester unis et solidaires.

Table des matières
Introduction 1
Chapitre 1- The new animal phylogeny: Reliability and Implications 3
Partie A- Définitions et concepts
Chapitre 2- Description et comparaison de la formation des feuillets 8
embryonnaires chez les métazoaires
1. Cadre général du développement des métazoaires 9
1.1. La fécondation: créer un nouvel organisme 10
1.2. Le clivage: organiser la multicellularité 10
1.3. La gastrulation: établir les feuillets embryonnaires 13
1.4. L'organogénèse: réaliser le plan d'organisation 15
2. Développement précoce de quelques métazoaires 15
2.1. Les ecdysozoaires 15
2.1.1. Drosophila melanogaster (Arthropode) 15
2.1.2. Caenorhabditis elegans (Nematode) 17
2.2. Les lophotrochozoaires 19
2.2.1. Patella vulgata (Mollusque) 19
2.3. Les deuterostomiens 23
2.3.1. Paracentrotus lividus (Echinoderme) 23
2.3.2. Halocynthia roretzi (Urochordé) 26
2.3.3. Branchistoma floridae (Céphalochordé) 27
2.3.4. Danio rerio (Vertébré) 29
2.3.5. Xenopus laevis (Vertébré) 31
2.3.6. Gallus gallus (Vertébré) 34
2.3.7. Mus musculus (Vertébré) 37
3. Remarques et conclusions 41
3.1. L'unité dans la diversité 41
3.2. De l'existence et de l'homologie des feuillets embryonnaires 42
3.3. Conclusions 44
Chapitre 3- Comprendre les mécanismes génétiques gouvernant 45
la formation du mésoderme
1. La formation du mésoderme: quelques généralités 46
2. Contrôle génétique de la formation du mésoderme 46
chez Drosophila melanogaster

2.1. Etablissement de l'axe Dorso-ventral chez D. melanogaster 46
2.2. La détermination du mésoderme 49
2.3. La différenciation précoce du mésoderme 51
3. Contrôle génétique de la formation du mésoderme chez les vertébrés 52
3.1. La formation du mésoderme chez Xenopus laevis 52
3.2. Le contrôle du mésoderme chez les autres vertébrés 56
4. Comparaison Ecdysozoaires / Deutérostomiens 58
5. Conclusions 60
Partie B- Approche expérimentale
Chapitre 4- Caractérisation de gènes homologues de Snail chez Patella vulgata 61
1. Choix d'un organisme modèle représentant les lophotrochozoaires 62
2. Un organisme modèle: Patella vulgata 63
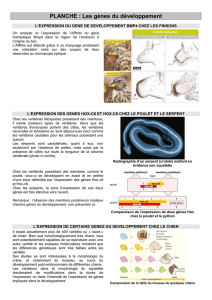
3. Gros plan sur le gène Snail 64
4. Recherche de gènes homologues de Snail 68
4.1. L'approche par PCR 68
4.2. L'alignemment des séquences disponibles en 1996 69
4.3. Conception d'oligonucléotides spécifiques de Snail 72
4.4. Amplification d'un fragment Snail chez Patella vulgata 74
5. Crible de banques d'ADN complémentaires 74
6. Recherche de gènes paralogues par Southern Blot 77
6.1. Analyse par Southern Blot: conditions expérimentales 77
6.2. Analyse par Southern Blot: résultats 77
7. Crible d'une banque d'ADN génomique 78
8. La famille des gènes Snail: un premier bilan 81
8.1. Chez Patella vulgata 81
8.2. Chez les Métazoaires 83
8.2.1. Niveau de similitude 84
8.2.2. Phylogénie de la famille des gènes Snail 85
8.3. Conclusions 88
9. Détermination des profils d'expression des gènes pv-Slak 89
9.1. Mise au point du protocole d'Hybridation in situ chez Patella vulgata 89
9.2. Profil d'expression de pv-Slak1 89
9.3. Profil d'expression de pv-Slak2 90
9.4. L'approche par anti-corps 91
9.5.
Conclusions 91
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
1
/
152
100%