chapitre 1

CLASSIFICATION DES ESPECES
La théorie cellulaire a été émise au 18ème siècle :
« LA CELLULE EST L’UNITE STRUCTURALE ET FONCTIONNELLE
DE TOUS LES ETRES-VIVANTS »
PROCARYOTES
EUCARYOTES
Métazoaire – animal « cellules animales pluricellaires »
Métaphyte – végtal « cellules végétales pluricellaires »
Protozoaire – prophyte – protiste « cellelules animales unicellaires »
ARCHEES
BACTERIES
ANIMAUX
PLANTES
CHAMPIGNONS
PROTISTES

GENERALITES SUR LA CELLULE
La cellule est la plus petite unité capable de réaliser toutes les réactions nécessaires à la vie
« respiration, nutrition … » Elle représente donc la plus petite unité de la matière vivante.
Elle s’alimente, respire, communique avec d’autres cellules et se reproduit.
Il existe des êtres-vivants unicellaires « une cellule : exemple les bactéries » et des êtres-
vivants pluricellaires « plusieurs cellules : exemple l’Homme »
Les êtres-vivants présentent une grande diversité. Pourtant tous sont constitués de cellule.
Chez un même organisme, les cellules présentent des différences de forme, de taille et aussi
au niveau de leurs organites. Elles sont spécialisées dans une fonction. Selon celle-ci une
cellule peut être plus ou moins riche en organites cellulaires. Malgré ces différences, les
cellules ont une organisation commune.
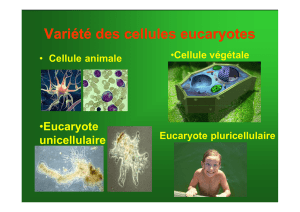
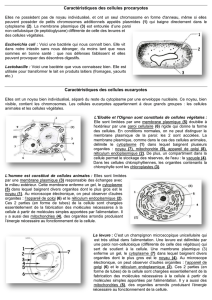
On distingue deux types de cellules : les cellules procaryotes et les cellules eucaryotes.
Les cellules eucaryotes possèdent un noyau. Une membrane plasmique délimite le
cytoplasme, dans lequel baignent les organites. Chaque organite exerce une fonction.
Parmi les cellules eucaryotes, les cellules végétales se distinguent des cellules animales par la
présence d’une paroi cellulaire et d’organites spécifiques comme une grande vacuole et des
chloroplastes.
Les cellules procaryotes, par exemple les bactéries, sont constituées d’un cytoplasme
dépourvu d’organites. L’information génétique est libre dans le cytoplasme.

ULSTRASTRURE D’UNE CELLULE EUCARYOTE ANIMALE

ULTRASRUCTURE D’UNE CELLULE EUCARYOTE VEGETALE

ULTRASRUCTURE D’UNE CELLULE PROCARYOTE
Un procaryote du latin pro « avant » et du grec caryon : « noyau » est un être-vivant dont la
structure cellulaire ne comporte ni de noyau - ni d’organites. Ces organismes se multiplient
par scissiparité « reproduction asexuée ». Les bactéries sont des procaryotes.
Il existe deux sortes de bactéries : les bactéries à gram+ et les bactéries à gram- qui sont
mises en évidence par une technique de coloration appelée coloration de Gram. Les
bactéries à Gram positif apparaissent mauves et bactéries à Gram négatif apparaissent roses
au microscope. La technique de coloration repose sur les caractéristiques membranaires et
de la paroi de la bactérie.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
1
/
25
100%