Contribution-a-une-etude- phytoecologiques.pdf

10
R
RE
EP
PU
UB
BL
LI
IQ
QU
UE
E
A
AL
LG
GE
ER
RI
IE
EN
NN
NE
E
D
DE
EM
MO
OC
CR
RA
AT
TI
IQ
QU
UE
E
E
ET
T
P
PO
OP
PU
UL
LA
AI
IR
RE
E
M
MI
IN
NI
IS
ST
TE
ER
RE
E
D
DE
E
L
L’
’E
EN
NS
SE
EI
IG
GN
NE
EM
ME
EN
NT
T
S
SU
UP
PE
ER
RI
IE
EU
UR
R
E
ET
T
D
DE
E
L
LA
A
R
RE
EC
CH
HE
ER
RC
CH
HE
E
S
SC
CI
IE
EN
NT
TI
IF
FI
IQ
QU
UE
E
U
UN
NI
IV
VE
ER
RS
SI
IT
TE
E
D
DE
E
T
TL
LE
EM
MC
CE
EN
N
F
Fa
ac
cu
ul
lt
té
é
d
de
es
s
S
Sc
ci
ie
en
nc
ce
es
s
d
de
e
l
la
a
N
Na
at
tu
ur
re
e
e
et
t
d
de
e
l
la
a
V
Vi
ie
e
e
et
t
d
de
es
s
S
Sc
ci
ie
en
nc
ce
es
s
d
de
e
l
la
a
T
Te
er
rr
re
e
e
et
t
d
de
e
l
l’
’U
Un
ni
iv
ve
er
rs
s
Département d’Ecologie et Environnement
Laboratoire de recherche
Ecologie et Gestion des Ecosystèmes Naturels
MEMOIRE
Présenté par :
CHERIF ISMAHENE
En vue de l’obtention du
Diplôme de Magister
En Ecologie et Biodiversité des Ecosystèmes Continentaux
Thème
Contribution à une étude phytoécologiques des
groupements à Tetraclinis articulata du littoral
de Honaine (Algérie occidentale).
Soutenu le……………………….., devant le jury composé de :
Présidente BELAIDI Nouria Professeur Université de Tlemcen
Encadreur BOUAZZA Mohamed Professeur Université de Tlemcen
Examinateurs LETREUCH-BELAROUCI Noureddine Professeur Université de lemcen
HASNAOUI OKKACHA M.C.A Université de Saïda
Invitée FELIDJ Manel M.A.A Université de Tlemcen
Année Universitaire 2011-2012

11
Dédicaces
A
mes très chers parents, qui sont toujours soucieux de
ma réussite, qu’ils trouvent ici le fruit de leurs sacrifices.
A
mes frères Djamel eddine, Abdel aziz, et Mohamed el
amine.
A ma très chère tante Hallouma, pour son soutien
permanent, ses encouragements et ses précieux conseils.
A
mon, très cher époux Houari, pour ses encouragements
aux moments difficiles, ses sacrifices, et sa confiance.
A
toute la famille Cherif et Sabri.
A
tous ce qui m’ont apporté d’aide de près ou de loin.
A
touts les étudiants de ma promotion et à tous mes amis.
Ismahene

12
Remerciements
Au terme de ce travail, il m'est agréable de remercier vivement tous ceux qui, grâce
à leur aide précieuse, ont permis la réalisation de ce travail.
Je dois remercier particulièrement Monsieur Bouazza Mohammed ; Professeur à la
Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers, de
l’Université Abou Bakr BELKAID – Tlemcen ; pour avoir accepté de diriger avec
beaucoup d’attention et de soins ce mémoire. Je lui suis très reconnaissante pour sa
disponibilité, sa bienveillance et son soutien permanent, et d’avoir prêté un intérêt constant
au sujet du mémoire. Je lui dois beaucoup pour le contenu du travail présenté, pour ses
critiques constructives et son aide aux différentes entraves rencontrées, pour sa gentillesse
et ses qualités humaines, ainsi que ses encouragements et sa confiance. Ces quelques mots
ne suffisent pas pour lui exprimer toute ma reconnaissance.
J’adresse ma gratitude à Mme Belaidi Nouria ; Professeur à la Faculté des Sciences
de la
Nature et de la Vie, des Sciences de la Terre et de l’Univers, de l’Université Abou Bakr
BELKAID –Tlemcen ; d’avoir accepté de me faire l’honneur de présider ce jury.
Je voudrais également remercier Monsieur Letreuch-Belarouci N. ; Professeur à la
Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers, de
’Université Abou Bakr BELKAID – Tlemcen ; pour avoir accepté de juger ce travail
Je remercie également Monsieur Hasnaoui Okkacha, Docteur et chargé de cours à
la Faculté des Sciences de l’Université de Saïda, pour son aide lors des sorties sur le terrain
et pour avoir accepté d’examiner ce travail et participer au jury de ce mémoire.
Je remercie également Melle Felidj Manel, chargé de cours à la Faculté des
Sciences de la
Nature et de la Vie, des Sciences de la Terre et de l’Univers, de l’Université Abou Bakr
BELKAID – Tlemcen ; pour avoir accepté mon invitation à ce jury.
Mes sincères remerciements sont adressés aussi à :
Monsieur Benabadji Noury ; Professeur à la Faculté des Sciences de la Nature et de
la Vie, des Sciences de la Terre et de l’Univers, de l’Université Abou Bakr BELKAID –
Tlemcen ; pour ses conseils et son aide concernant l’analyse pédologique.
Madame Stambouli-Meziane Hassiba, Docteur et chargé de cours à la Faculté des
Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers, de l’Université
Abou Bakr BELKAID – Tlemcen ; pour sa gentillesse et pour son aide concernant la
détermination des espèces et les traitements statistiques.
Monsieur Benzaim Ahmed, cartographe, pour son aide si précieuse en cartographie.
A tous ceux et celles qui ont participé de près ou de loin à l'élaboration de ce travail,
qu'ils trouvent ici ma haute considération.

13
ـــــا
ةرا ل ء هو ،اا ب ل ا ا ارد .
ت ت ـا ـا إ ارـا ه ف)را (Tetraclinis articulata ا
،اآ أ ، تا ا ا ارا آ ، ا
ا ت طا لا ىدأ آ اه فواو إ إ ، ،أو نا ف
او ا ه باا إ ا ا.
إ ارا ه رإ لا ا ا ل 14 ت إ ، ع
را Tetraclinis articulata 05 و و ت:Lavandula dentata 60 % ,
Clycotome intermedia 58 %, Pistacia letiscus 50 %, Globularia alypum 44%, Cistus
monspeliensis 44%.
ةرذ ىأو :Ulex europeus 38 %, Phyllirea angustifolia 28 %, Chamaerops humilis 32
%, Helianthemum pilosum 22 %, Urginea maritime et Asteriscus maritimus 36 %,
Ampelodesmamauritanicum26 %,Schismus barbatus 35 %.
ه ةر آأ عوvicia vilolsa 06% .
آ)(AFC آا ا اا ف ا ٌأو ت ارد
را ت ا. Tetraclinis articulata
حواـ ا ـا ةدز ىأ إ لـا نأ ا اه او
dématoralistaion إthérophytisation
ا تا :
تا أ- را)tetraclinisarticulata (- ا ا - تا – ن - ناهو– اا.

14
RÉSUMÉ
La zone sur laquelle porte notre contribution est localisées dans la partie occidentale
du Nord-Ouest Algérien, elle fait partie intégrante des monts des Traras.
Cette étude est consacrée à l’analyse phytoécologique des groupements à Tetraclinis
articulata du littoral de Honaine.
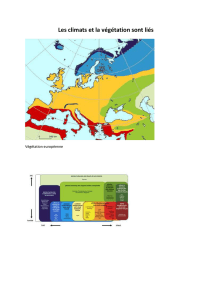
L’étude bioclimatique a révélé un décalage de certaines stations météorologiques, vers
des ambiances plus sèches sur le climagramme pluviothermique d’Emberger, témoignant ainsi
une tendance générale à l’aridité.
L’utilisation abusive des formations végétales, par l’homme et ses troupeaux, conduit,
selon la qualité des conditions édapho-climatiques, à différents stades de dégradation allant de
la simple altération partielle de la composition floristique à une perturbation totale.
Nous avons pu décrire une liste de 14 espèces dites caractéristiques du Tetraclinis
articulata ou encore les espèces qui constituent les groupements de cette dernière. Parmi eux
nous avons cinq espèces fréquentes, ou espèces constantes : Lavandula dentata 60 %,
Clycotome intermedia 58 %, Pistacia letiscus 50 %, Globularia alypum 44%, Cistus
monspeliensis 44%. Parmi les espèces caractéristiques du Tetraclinis articulata, il y a des
espèces rares dont la fréquence ne dépassant pas les 40%, parmi eux : Ulex europeus 38 %,
Phyllirea angustifolia 28 %, Chamaerops humilis 32 %, Helianthemum pilosum 22 %,
Urginea maritime et Asteriscus maritimus 36 %, Ampelodesma mauritanicum 26 %,
Schismus barbatus 35 %. L’espèce la plus rare est Vicia villosa avec 6 %.
L’analyse factorielle des correspondances nous a permis une étude globale très
synthétique des données, et de traiter conjointement les variables floristiques et écologiques.
Nous avons conclu après cette analyse que le passage d’un groupement à un autre répond au
déclenchement des processus de désertification dont les modalités diffèrent ; allant de la
dématorralisation jusqu'à la thérophytisation
Mots clés : Échantillonnage - Tetraclinis articulata – Phytoécologie – Botanique – Tlemcen –
Oranie – Algérie.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
1
/
207
100%