TP2

TP2
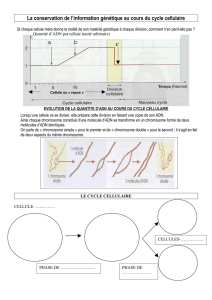
la duplication de l’ADN nécessite une réplication
1. Le problème :
Depuis Watson et Crick (1953), on sait que l'ADN est une molécule formée de deux brins complémentaires,
formant une double hélice. Voir : http://librairiedemolecules.education.fr/outils/adn/#top
Dès leur publication originale sur la structure de
l'ADN, Watson et Crick ont proposé que cette
double hélice puisse s'ouvrir, permettant ainsi la
synthèse de nouveaux brins, complémentaires
des brins originaux.
L'ADN peut ainsi servir de matrice à sa propre
réplication, étape essentielle du cycle cellulaire.
Cette duplication de l'ADN (et donc des
chromatides) permet de passer de chromosomes à une chromatide à des chromosomes possédant deux
chromatides identiques, portant la même information génétique. Lors de la mitose, ces deux chromatides
sont réparties, chaque cellule - fille héritant d'une chromatide de chaque chromosome. On obtient ainsi
deux cellules possédant la même information génétique que la cellule - mère.
Le problème qui se posait à Meselson et Stahl était alors de comprendre comment se réalisait cette
réplication : selon quelles modalités passe-t-on d'une molécule d'ADN formée de deux brins à deux
molécules d'ADN bicaténaires identiques ?
2. Les hypothèses
Pour expliquer la duplication d'un ADN bicaténaire, trois modèles ont été proposés. Ces modèles se basent
tous sur l'utilisation de la molécule d'ADN "mère" comme matrice pour sa réplication, mais selon des
modalités différentes :
• Schématisez les 3 hypothèses en utilisant les couleurs indiquées
Hypothèse 1: modèle conservatif
A partir d'une molécule d'ADN
bicaténaire "mère", on forme une
nouvelle molécule d'ADN
bicaténaire.
On garde donc ici une molécule
"mère", non modifiée (elle est
donc conservée), tout en "créant"
une nouvelle molécule ("fille").
Hypothèse 2: modèle semi-
conservatif
On dissocie les deux brins de la
molécule d'ADN bicaténaire "mère".
Chaque brin sert donc de matrice à la
synthèse d'un brin complémentaire,
l'ensemble reformant une molécule
d'ADN bicaténaire. Chaque nouvelle
molécule "fille" ne conserve donc que
la moitié de la molécule "mère".
Hypothèse 3: modèle
dispersif
On ne conserve aucun brin
intact.
La copie se réalise par
fragments dispersés dans
l'ensemble de l'ADN,
permettant de former les
deux molécules d'ADN
bicaténaires "filles".
Légendes :
Couleur rouge : Molécule d'ADN "mère" et son devenir.
Couleur bleue : ADN néo-formé.

• Vérifiez à l’échelle de l’ADN:
http://pedagogie.actoulouse.fr/svt/serveur/lycee/gueraut/stahl/3exphyp3d.htm
• Vérifiez à l’échelle des nucléotides :
http://pedagogie.ac-toulouse.fr/svt/serveur/lycee/gueraut/stahl/5exphypnucl.htm
APPELEZ LE PROFESSEUR
3. L’expérience : http://www.conceptcours.fr/www/prem_s/genet/doc_gene/mesel_stahl_exp.swf
Les expériences sont réalisées sur la bactérie E-Coli, qui se multiplie activement quand elle est cultivée sur
un milieu favorable et réplique alors intensément son ADN.
Les bactéries sont cultivées dans deux milieux différents : L’un contient des nucléotides « légers » intégrant
de l’azote « léger » 14N. L’autre contient des nucléotides « lourds » intégrant de l’azote « lourd » 15N.
Des bactéries, d’abord cultivées depuis de nombreuses générations sur un milieu contenant des nucléotides
15N, sont prélevées et transférées sur un milieu normal à nucléotides 14N.
A chaque étape, l’ADN des bactéries en culture est extrait et centrifugé à grande vitesse dans un tube
contenant une solution de densité appropriée. Après centrifugation, l’ADN se stabilise ainsi à un niveau
correspondant à sa densité.
• Comprendre l’expérience :
Résultats
densité
Constitution de
l’ADN
Etapes de
l’expérience
Résultats
Déductions
1,724
1. Culture sur 15N
(n cycles cellulaires)
1,710
2. Culture sur 14N
(1 cycle)
1,717
3. Culture sur N14
(2cycles)

• Confronter le résultat 2 aux trois hypothèses. Laquelle pouvez-vous éliminer ?
• Confronter le résultat 3 aux hypothèses restantes. Laquelle pouvez-vous éliminer ?
• Vérifier : http://pedagogie.ac-toulouse.fr/svt/serveur/lycee/gueraut/stahl/6expcorr.html
APPELEZ LE PROFESSEUR
Meselson et Stahl ont complété leur travaux :
http://www.conceptcours.fr/www/prem_s/genet/doc_gene/con20ani.swf
NB : le centromère correspond à une zone du chromosome où la réplication est tardive et n’intervient qu’à l’anaphase.
• Réalisez un schéma du devenir d’un molécule d’ADN au cours de la mitose (conservez les codes
couleurs du début)
Phase G1
Phase S
Fin de phase S
Prophase
Métaphase
Chromosome :
Chromosome :
Chromosome
Chromosome
Bilan :
Anaphase
télophase
Chromosome
Chromosome
Je vérifie que j’ai compris :
https://www.education-et-numerique.fr/0.3/activity/embed.html?id=54d0d86d3361eb54776f19ad
!
1
/
3
100%