UE9 – Immunopathologie et Immunointervention Lefebvre d’Hellencourt

UE9 – Immunopathologie et Immunointervention
Lefebvre d’Hellencourt
Date : 10/03/2017 Plage horaire : 8h30-10h30
Promo : PCEM2 2015-2016 Enseignant : Dr Lefebvre d’Hellencourt
Ronéistes : BLANPAIN Amélie / MERMILLOD Aurore
Introduction à l’immunologie : 1ère partie
I – Présentation générale
1. La variole
2. Pasteur
3. Définitions
II – Le système immunitaire
III – L’infection
1. Les lymphocytes naïfs
2. Les cellules mémoires
IV – L’hématopoïèse
1. Les lymphocytes
2. Les monocytes
3. Les cellules dendritiques
4. Les neutrophiles
5. Les éosinophiles
6. Les basophiles
7. Les mastocytes
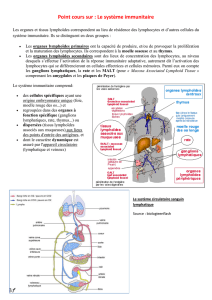
V - Les organes lymphoïdes
1.Les organes lymphoïdes primaires
2.Les organes lymphoïdes secondaires
VI- Les antigènes (1 er
partie)
1.Définition
2.Propriétés
3.Les différents types d’antigène
4.Valence
1

Le cours est basé sur le livre Immunobiologie, 5e édition (accessible en ligne sur le site du NCBI)
I – Présentation générale
Notions historiques :
L'immunologie reste un domaine assez récent, bien que les concepts soient très anciens. Dans les années
1900, il y a eu une évolution des théories avec la théorie humorale et la théorie cellulaire (humorale : théorie
des anticorps et des lymphocytes tandis que la théorie cellulaire fait intervenir la phagocytose, les
lymphocytes T ainsi que les notions de soi), deux théories qui ont finalement pu s’associer mais qui, au
départ, étaient opposées. Les notions de système intégré ont permis de replacer le système immunitaire dans
l’ensemble des systèmes, à toute la physiologie.
Il y a eu des progrès techniques ayant permis d'avancer dans la compréhension de l'immunologie avec des
techniques d'observations des tissus et des cellules qui ont beaucoup évolué avec l'apparition de la
microscopie, de même qu'avec les progrès importants en biochimie, génétique, physique nucléaire et en
biomoléculaire.
On a aussi connu des évolutions au niveau des concepts, avec des notions telles que la tolérance qui ont
permis de faire avancer les connaissances sur l’immunité. On est maintenant dans une ère dit post-
génomique car la plupart des génomes ont été séquencés (on a ainsi une vue globale de l’ensemble des gènes
quand on veut étudier un phénomène). Il existe maintenant des techniques parallèles de bioinformatique
permettant d’intégrer toutes ces données génomiques.
Deux grands exemples illustrent les progrès en immunologie…
1. La variole
Cette pathologie a provoqué des épidémies très importantes, bien qu'elle soit de nos jours
éradiquée.
Il y a eu des cas d'abord importants en Asie (début en Chine), puis il y a eu des migrations vers l'Amérique
du nord (où la population était non immunisée) avec des dégâts importants au niveau des populations. Cette
pathologie est due à un virus du groupe des POX Virus, qu'on retrouve dans les sérosités et transmis soit par
2

contact direct (voie cutanée ou muqueuse), soit indirect (voie respiratoire ou digestive). Le taux de mortalité
pouvait varier de 25 à 45% selon les souches. Les moyens présents pour lutter contre la variole étaient très
variables (inoculation, prières...).
Il y a plusieurs phases qui caractérisent la variole :
-l'incubation : elle dure de 8 à 10 jours (phase lente)
-l'invasion : avec des signes grippaux (frissons, température...)
-l'éruption : avec l'apparition de vésicules et pustules (visages en premier puis le reste du corps)
-la suppuration : 3 jours après la phase d'éruption
-la dessiccation qui permet l'assèchement des vésicules et ainsi la formation de croûtes qui tombent
laissant des cicatrices indélébiles. (pour les patients qui survivent)
Lorsque l'on parle de la variole, cela concerne à la fois les personnes atteintes qui vont mourir de
l'infection et les personnes qui vont survivre mais être défigurées.
Parmi les techniques utilisées pour lutter contre la variole, on a d’abord eu la variolisation. Cette
technique consistait à prélever du pus de patient atteint de variole puis à l'injecter à des patients sains
pour essayer de les protéger. Il y avait des résultats intéressants car cela protégeait les personnes mais il y
avait tout de même une mortalité de 2%, ce qui était important, et un risque de propagation de l'épidémie.
Cette technique fonctionne car la mortalité par variole était de 40 à 50% mais pour les 2% qui ne
survivaient pas ce n’était pas suffisant.
Cette technique de variolisation a été importée de l’Asie vers l’Europe par une femme de diplomate anglais,
atteinte de la variole donc très sensible à ce problème. Elle fut d’abord appliquée sur des prisonniers anglais
puis plus largement sur la population.
Ce n'est que plus tard que la protection fut optimale grâce à Edward Jenner en 1796, avec l'apparition de la
vaccination en ayant comparé la variole humaine avec la variole animale (similitudes entre l'homme et la
vache). Il a utilisé des vaches infectées pour essayer de voir si cela pouvait protéger des hommes ou pas. Il a
injecté à un enfant de 7 ans sain du pus de la variole de vache ce qui a permis de le protéger.
La variole chez la vache est la vaccine, cowPOX virus : ainsi la vaccine injectée à un homme protège de la
variole humaine, d'où le nom de vaccin, puis « vaccination » qui a été ensuite utilisé. L’avantage par rapport
à la variolisation est un taux de mortalité quasiment nul.
La vaccination a permis assez rapidement d'éradiquer la variole (quand elle a été enfin utilisée de façon
générale) grâce aux campagnes mondiales de vaccination qui ont commencé dans les années 60 (1977 :
denier cas de variole qui est apparu en Étiopie). C'est l'exemple que l'immunologie peut combattre une
épidémie dramatique.
Actuellement, la variole n'est plus une menace ; il ne reste que quelques souches de ce virus présents dans
des laboratoires de recherche aux USA et en Russie (approche de protection contre-terrorisme...)
3

2. Pasteur
Au cours de sa vie, il a eu plusieurs carrières :
- Chimiste (travaux sur la dissymétrie moléculaire)
-Microbiologiste avec la fermentation : il est à l'origine de beaucoup de techniques de
microbiologie. Par exemple, il a permis de démontrer que la génération spontanée n'existait pas (à l'époque
on pensait que si on avait une solution riche en nutriments, l'apparition de micro-organismes était spontanée,
sans explication). Il a montré que si on utilisait un bécher avec un bouchon filtrant, il empêchait l'apparition
de la génération de microbes. Cela a eu des conséquences importantes, notamment en matière d’asepsie.
-L'agronomie : il a travaillé sur les maladies du vin et de la bière (problèmes importants pour les
industries de France de l'époque, par rapport aux parasites qui étaient dans le vin, à la fabrication de la
bière…) Il a développé la pasteurisation, qui a permis de conserver de façon durable des aliments et des
solutions.
-La maladie des vers à soie qui était une maladie importante de l'époque dans l'agroalimentaire
-Travaux sur les maladies infectieuses avec la découverte des staphylocoques, streptocoques et
pneumocoques.
-Travaux sur les méthodes d'atténuation de la virulence des microbes, afin de procéder ensuite à
des vaccinations (choléra). Ce qui a rendu célèbre Pasteur, ce sont ses travaux de 1985 sur la rage : par sa
méthode d'atténuation, il a injecté des souches atténuées à un patient atteint (enfant de 9 ans), il a pu le
sauver, ce qui a été un scoop médiatique, permettant de récolter des fonds et de fonder l'Institut Pasteur.
3. Définitions
L'immunité : c'est l'ensemble des réactions qui vont tendre à éliminer des substances étrangères (par
exemple : virus, bactéries, champignons, parasites, macromolécules, cellules animales étrangères, cellules
étrangères de la même espèce). Cela va sous-tendre aux notions de soi et de non soi.
L'immunologie : c'est une discipline biologique et médicale qui va étudier les processus de reconnaissance
de substance antigénique.
La réponse immunitaire : c'est l'ensemble des conséquences de la reconnaissance des substances
antigéniques.
Le système immunitaire : C'est un système complexe de facteurs humoraux et d'interactions cellulaires qui
vont permettre la réponse immunitaire.
La vaccination : méthode de stimulation de l’immunité vis-à-vis d’une infection grâce à l’injection d’un
microorganisme ou un de ses produits sous forme inoffensive.
4

II – Le système immunitaire
Le système immunitaire va assurer 3 fonctions fondamentales :
- la définition de l'individu (le soi)
-la reconnaissance de signaux moléculaires étrangers (le non soi)
- l'organisation de la défense du soi
La définition de l'individu fait appel aux notions de soi et non soi. Or pour reconnaître ce qui
n'est pas du soi, il faut reconnaître des signaux moléculaires des substances étrangères et ainsi
le système immunitaire pourra organiser la défense.
Dans l'immunopathologie, il y aura le dérèglement du système immunitaire :
- un déficit de la réponse immunitaire (exemple : le sida, dans certains types de cancers)
- l'auto-immunité (exemple : le lupus, sclérose en plaque)
-hypersensibilité (exemple : l'allergie ou l'asthme)
III – L’infection
L'infection est due à un pathogène. Pour avoir une infection, il faut que le pathogène franchisse
différentes barrières comme la barrière physique qui peut être la peau ou les muqueuses (parfois
fragilisées). Si le microbe pénètre dans l'organisme, des systèmes de défense vont se mettre en place :
-La 1ere défense : l'immunité innée, qui peut être suffisante pour éliminer la prolifération du
microbe et ainsi l'élimination du pathogène. Elle consiste en une réaction immunitaire et une réaction
inflammatoire. Si l'immunité innée est insuffisante, on aura ensuite la mise de place de l'immunité
spécifique.
-La 2ème défense : l'immunité spécifique fait intervenir différentes cellules qui vont permettre de
combattre ce pathogène. C'est le cas de cellules capables de capter l'antigène et de l'emmener dans un
organe lymphoïde secondaire où elles pourront interagir avec d'autres cellules (les lymphocytes) qui vont
reconnaître l'antigène capté.
L'interaction entre les cellules présentatrices de l'antigène et les lymphocytes va permettre d'activer
ces lymphocytes et les rendre effecteurs (à la base ce sont des lymphocytes naïfs).
Ces lymphocytes effecteurs vont avoir 2 évolutions :
-combattre le pathogène et l’éliminer (une fois l'élimination du pathogène faite, les lymphocytes
meurent par apoptose)
- la mise en place des cellules mémoires (une petite population)
1. Les lymphocytes naïfs
Ce sont eux qui vont aller à la rencontre des cellules présentatrices de l'Ag et qui vont ensuite être activés en
effecteurs. Il faut que ces lymphocytes naïfs aient :
-la capacité de reconnaître l'Ag, avec une diversité importante ; soit une diversité de récepteurs ++
- une sélection performante pour que ces lymphocytes naïfs ne reconnaissent pas le soi (avec l’élimination
des lymphocytes auto-réactifs).
Les lymphocytes naïfs doivent donc reconnaître le non soi de façon suffisamment large pour pouvoir
prendre en compte l'ensemble des pathogènes possibles.
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
1
/
22
100%