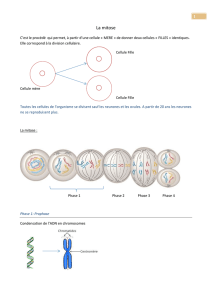
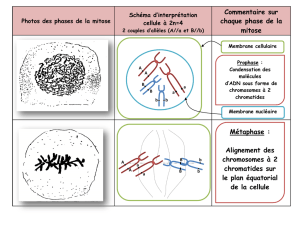
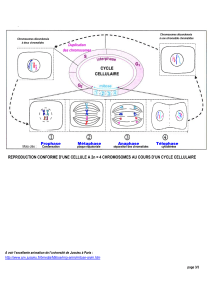
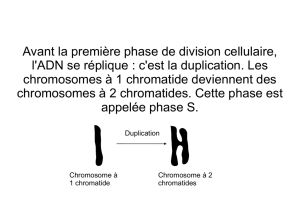
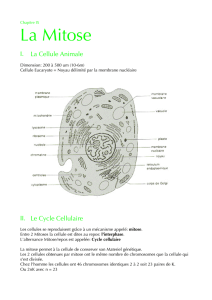
Chapitre 1 : Unicité chimique des êtres vivants

Chapitre 1 :
Unicité chimique des êtres vivants
1) Associez le nom de chacun de ces lipides à sa formule chimique. Lesquels sont présents dans
les membranes plasmiques (voir Chapitre 3) ? Comment définir un lipide ?
Conseil : Repérez les structures caractéristiques des différents types de lipides.
a) Triglycéride
b) Cortisol (sachant qu’il dérive du cholestérol)
c) Acide gras
d) Cholestérol
e) Phosphatidylinositol
I) II)
III)
IV) V)
2) En sachant que l’acide aminé glycine correspond à l’extrémité C-terminale, représentez en
détail la structure du peptide suivant, en indiquant notamment les liaisons peptidiques.
Peptide : Gly–Ala–Cys
Conseil : Souvenez-vous de la réaction de formation d’un peptide à partir de plusieurs acides aminés.

Réponses :
1)
a) Triglycéride : formule III
- Non présent dans la membrane plasmique.
- Hydrolysables, les triglycérides constituent une réserve d'énergie grâce aux acides gras qu’ils
contiennent. Ces derniers sont dégradables par la β-oxydation et fournissent, via la chaîne
respiratoire, de l’énergie à la cellule. Ils sont stockés dans les cellules adipeuses.
b) Cortisol : formule IV
- Non présent dans la membrane plasmique.
- Le cortisol est une hormone stéroïde sécrétée par le cortex de la glande
surrénale et fabriquée à partir de cholestérol. Il permet de réguler le
métabolisme des glucides (néoglucogenèse), des graisses et des protéines.
c) Acide Gras : formule V
- Comme tel, non présent dans la membrane plasmique.
Par contre, les phospholipides, qui contiennent des acides gras, sont bien présents dans la
membrane plasmique (voir Chapitre 3).
- Les acides gras constituent une importante réserve d’énergie contenue dans la cellule. Ils sont
stockés sous la forme de triglycérides dans des gouttelettes lipidiques.
d) Cholestérol : formule I
- Présent dans la membrane plasmique (voir Chapitre 3).
- Le cholestérol est un composant majeur des membranes cellulaires
animales. Il contribue à leur stabilité et au maintien de leurs structures en
s'intercalant entre les phospholipides.
e) Phosphatidylinositol : formule II
- Présent dans la membrane plasmique (voir Chapitre 3).
- Les phospholipides, avec les glycolipides et les stérols, sont les
constituants essentiels des membranes cellulaires. La nature des
acides gras des phospholipides joue un rôle important dans la fluidité
des membranes, ainsi que dans la perméabilité à certaines molécules
ou à certains ions.

Parmi ces lipides, les seuls qui soient présents dans la membrane plasmique sont donc le cholestérol
et le phosphatidylinositol (voir Chapitre 3).
Une définition possible les lipides : Classe hétérogène de molécules organiques insolubles dans
l’eau, solubles dans les solvants organiques apolaires (benzène, chloroforme, éther…) et
caractérisées par la présence dans la molécule d’au moins un acide gras. Cette dernière
caractéristique n’est toutefois pas présente chez les stéroïdes. Ceux-ci, insolubles dans l’eau, sont
tout de même rattachés aux lipides, mais on parlera plutôt de « composés à caractère lipidique ».
À vrai dire, il n'existe pas vraiment de définition unique reconnue par la communauté scientifique, les
« lipides » désignant un ensemble de molécules aux structures et aux fonctions extrêmement
variées.
2) Par convention, le premier acide aminé de la structure primaire d’un peptide est celui dont la
fonction amine n’est pas engagée dans une liaison peptidique, constituant l’extrémité NH2-
terminale du peptide. Ce peptide est donc formé de trois acides aminés :
HOOC–GlycineAlanineCystéine–NH2 ou H2N–CystéineAlanineGlycine–COOH
Deux liaisons peptidiques (CO–NH) sont présentes dans ce peptide et sont entourées ci-dessous :
Cystéine
Alanine
Glycine
Cystéine
Alanine
Glycine
= Liaison peptidique

Chapitre 2 :
Unicité structurale du vivant
et méthodes d’étude de la cellule
1) Pour chacun des organismes/organites/constituants suivants, citez le moyen (œil humain,
microscope optique ou microscope électronique) que vous utiliseriez pour le visualiser :
a) l’ADN
b) une mitochondrie
c) un œuf humain
d) une cellule eucaryote
e) une paramécie
f) une cellule nerveuse
g) des ribosomes
h) un virus
Conseil : Établissez une échelle de grandeur pour chaque élément à observer.
2) Il existe de nombreux colorants permettant d’identifier la cellule et sa constitution lors de
son observation au microscope. La coloration peut être :
a) Simple : utilisation d’un colorant unique
b) Combinée : utilisation de plusieurs colorants
Pour chacun des colorants suivants :
- vert de méthyle-pyronine
- réactif de Schiff
- hématoxyline/éosine
Pouvez-vous préciser s’il s’agit d’une coloration simple ou combinée et indiquer ce qu’ils
mettent en évidence ?
Conseil : L’utilisation de plusieurs colorants permet d’ajouter du contraste entre différents éléments
dans la préparation à observer au microscope.
Réponses :
1) Le choix de l’instrument d’observation dépend de la taille de l’objet à examiner, mais aussi du
type d’informations que l’on souhaite retirer de l’analyse. L’œil humain ne peut distinguer des
objets en dessous de 100 µm. Les pouvoirs de résolution des microscopes optique et
électronique sont respectivement de 200 nm et 0,2 nm.

a) Microscopie électronique. L’ADN est une molécule constituée de 2 chaînes nucléotidiques
enroulées en une double hélice (voir Chapitre 1). Son diamètre est de 2 nm, il ne peut donc être
observé qu’au microscope électronique.
b) Microscope optique et microscope électronique. Une mitochondrie est un organite des
cellules eucaryotes dont la dimension est de 1 à 10 µm de long et 0,5 à 1 µm de diamètre (voir
Chapitre 9). Bien que les mitochondries soient déjà visibles en microscopie optique, en
particulier à l’aide d’un microscope à contraste de phase, l’examen de leur organisation
structurale interne nécessite l’emploi d’un microscope électronique.
c) Microscope optique et microscope électronique. La taille de l’œuf humain (100 µm de
diamètre) permet difficilement de l’observer à l’œil nu. Observer un œuf humain nécessite donc
un microscope optique ou un microscope électronique pour les détails plus fins.
d) Microscope optique, œil humain et microscope électronique. La taille moyenne d’une cellule
eucaryote varie entre 10 et 100 µm de diamètre. En conséquence, à l’exception de quelques
cellules de grande taille comme l’œuf de poule qui sont visibles à l’œil nu, les cellules eucaryotes
seront observées à l’aide d’un microscope optique. L’organisation structurale interne de ces
cellules pourra être étudiée en microscopie électronique.
e) Microscope optique, microscope électronique et œil humain. Étant donné que la paramécie
est un protiste cilié dont la taille varie de 50 à 300 µm de long suivant les espèces, cet
unicellulaire peut, dans la majorité des cas être, observé à l’aide d’un microscope optique. Pour
les espèces de grande taille, elles pourront être observées à l’œil nu. Pour l’examen de leur
structure interne, il faudra le microscope électronique.
f) Microscope optique, œil humain et microscope électronique. La taille des neurones variant
entre 5 et 120 µm de diamètre, leur observation sera réalisée au moyen d’un microscope
optique. Cependant, les invertébrés peuvent présenter des cellules nerveuses de plus grande
taille, visibles à l’œil nu. Par exemple dans le ganglion viscéral de l’aplysie (lièvre de mer,
mollusque gastéropode sans coquille), un neurone peut atteindre 1 mm de diamètre. Le
microscope électronique sera nécessaire pour l’analyse de la structure interne des neurones.
g) Microscope électronique. Les ribosomes sont des structures subcellulaires de 25-30 nm de
diamètre qui requièrent le microscope électronique pour leur observation (voir Chapitre 5).
h) Microscope électronique. Les virus se caractérisant par leur très petite taille (< 250 nm), le
microscope électronique est indispensable à leur observation. Cependant, il a été récemment
découvert des virus géants (400 nm) qui peuvent être observés en microscopie optique.
2) La coloration par le vert de méthyle-pyronine est une méthode cytochimique utilisant 2
colorants (coloration combinée, b) pour différencier l’ADN de l’ARN. La chromatine, riche en
ADN, est colorée en vert par le vert de méthyle tandis que le nucléole et les régions
cytoplasmiques avec un abondant contenu en ribosomes, structures riches en ARN, sont colorés
en rouge par la pyronine.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
1
/
39
100%