Télécharger le fichier

1
Chapitre 1 : La cellule acineuse pancréatique
La cellule acineuse pancréatique exocrine est une cellule eucaryote spécialisée dans la
sécrétion de proenzymes inactives, les zymogènes. Cette cellule appartient à un tissu
spécialisé qui exerce sa fonction spécifique au sein d’un organe, le pancréas. Toutes les
cellules d’un tissu sont physiquement et fonctionnellement liées les unes aux autres. Leur
fonctionnement est coordonné par voie nerveuse et/ou humorale.
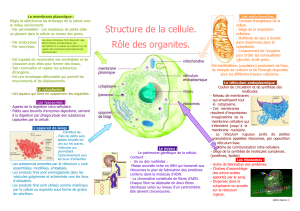
En coupe, la cellule acineuse pancréatique se présente sous forme trapézoïdale ; elle possède
un gros noyau en position basale et une très forte compartimentation liée à l’existence d’un
réseau membranaire, le réticulum endoplasmique, qui se développe à partir de la
membrane nucléaire externe. Ce réticulum est recouvert de très nombreux ribosomes
(réticulum endoplasmique rugueux, RER) qui sont le siège d’une importante synthèse
protéique. Le réticulum non recouvert de ribosomes (réticulum endoplasmique lisse) est le
siège de la synthèse des lipides. Ces deux réticulums semblent en continuité.
Les ribosomes accrochés au RER transloquent les chaînes polypeptidiques qu’ils
synthétisent dans la lumière du réticulum où elles passent par une maturation qui se poursuit
dans les dictyosomes avant d’être excrétées, dirigées vers des organites ou inclus dans la
membrane plasmique. Les ribosomes libres synthétisent les protéines hyaloplasmiques.
Toutes les membranes qui délimitent les compartiments cellulaires et la membrane plasmique
sont constituées d’une bicouche lipidique dans laquelle les protéines diffusent latéralement.
Ces membranes ont les propriétés d’une mosaïque fluide dont les composants se déplacent
avec plus ou moins de liberté les uns par rapport aux autres.
La cellule acineuse pancréatique est polarisée, la répartition de ses organites n’est pas
aléatoire ainsi que celle des protéines transmembranaires qui demeurent dans certaines
régions par des jonctions étanches. La partie basale de la cellule renferme le réticulum
endoplasmique, les ribosomes et de nombreuses mitochondries, signes d’une importante
activité de synthèse protéique. Au-dessus du noyau se situe l’appareil de Golgi spécialisé
dans la distribution des différentes vésicules dans les différentes régions de la cellule.
Cette cellule prélève du côté basal les nutriments (oses, acides aminés, acides gras) l’eau, le
dioxygène et les ions dont elle a besoin pour son métabolisme. Avec l’ATP qu’elle produit
principalement dans les mitochondries et son information génétique, la cellule acineuse
pancréatique synthétise des protéines pour son propre compte (renouvellement moléculaire)
mais aussi pour l’ensemble de l’organisme.
Le maintien de la forme, de l’ultrastructure de la cellule et le déplacement des organites est
assuré par le cytosquelette qui est constitué de trois types d’édifices supramoléculaires : les
filaments intermédiaires, les filaments fins et les microtubules.
Les filaments intermédiaires (kératine) sont des structures statiques qui participent au
maintien de la forme de la cellule et assurent la liaison mécanique entre les cellules par
l’intermédiaire des desmosomes.
Les filaments fins d’actine sont le plus souvent labiles, ils sont plutôt situés principalement
sous la membrane plasmique où ils participent à la formation des jonctions adhérentes. Ils
sont souvent organisés en faisceaux et interviennent dans le maintien de la forme de la
cellule. Ils interagissent avec de très nombreuses protéines dont des moteurs moléculaires
comme les myosines. Actine et myosine forment le système contractile de la cellule. La
dépolymérisation de l’actine est à l’origine de changement de pression osmotique utilisée
par la cellule pour se déformer ou se déplacer. Ils servent aussi de rails pour le transport
vésiculaire.
Les microtubules sont des structures permanentes des cils et des flagelles, ils constituent
aussi des rails sur lesquels circulent des organites par l’intermédiaire de moteurs
moléculaires (dynéines, kinésines). Durant les divisions cellulaires, ils se dépolymérisent et
leurs unités constitutives servent à construire l’appareil mitotique qui rayonne à partir des
centrioles.

2
La cellule acineuse est traversée par un flux de matière (prélèvement d’acides aminés dans le
milieu intérieur, sécrétion de protéines dans le milieu extérieur) qui est accompagné par un
flux membranaire qui augmente la surface de la membrane plasmique à la suite de
l’exocytose. Le maintien de la surface membranaire se réalise par différentes formes
d’endocytose du côté basal.
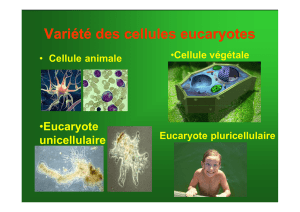
Chapitre 2 : La cellule du parenchyme palissadique
Les cellules du parenchyme palissadique sont des cellules foliaires spécialisées dans la
conversion de l’énergie lumineuse en énergie chimique et dans la fixation et la réduction
du dioxyde de carbone en matière organique. En réduisant le dioxyde de carbone, elles
réalisent la photosynthèse, elles sont donc autotrophes au carbone. Ces cellules présentent
de très nombreux organites communs avec la cellule animale (noyau, réticulum, Golgi et ses
dérivés, mitochondries, ribosomes), mais elles ne possèdent ni filaments intermédiaires, ni
centrioles. En revanche, elles se distinguent par la possession d’une paroi, d’une vacuole et
de chloroplastes.
Ces cellules sont approvisionnées par les stomates en dioxyde de carbone qui diffuse par la
suite dans les espaces intercellulaires. L’eau nécessaire à la photosynthèse arrive par le
xylème. Le saccharose produit par photosynthèse est exporté vers les cellules hétérotrophes
de la plante par le phloème. Ces deux tissus forment les faisceaux cribro-vasculaires. Les
nombreux méats et lacunes entre les cellules du parenchyme facilitent la circulation des gaz
entre l’atmosphère et les cellules.
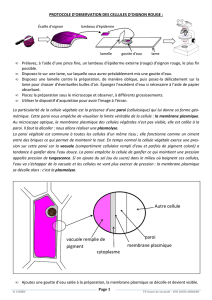
La cellule végétale possède une paroi pecto-cellulosique ou paroi squelettique. Il s’agit
d’une matrice extracellulaire qui définit un compartiment extracellulaire, l’apoplasme, dans
lequel circule une partie de l’eau et des ions prélevés dans le sol. La paroi pecto-cellulosique
est composée essentiellement de glucides (cellulose, hémicellulose, pectines) et de quelques
protéines. La paroi primaire, peu épaisse, est extensible sous l’effet de la pression de
turgescence exercée par la vacuole. La paroi secondaire qui se dépose entre la paroi primaire
et la membrane est plus épaisse et se forme au cours de la différenciation cellulaire. Son
dépôt marque la fin de la croissance de la cellule. La paroi n’est pas présente sur toute la
membrane. Il reste des zones qui en sont dépourvues, ce sont les plasmodesmes qui sont des
solutions de continuité entre les cellules adjacentes. Les cellules vivantes d’un végétal sont
donc en continuités membranaire et hyaloplasmique, elle forme un compartiment appelé le
symplasme.
Certaines cellules synthétisent en plus de la lignine qui se dépose dans la paroi où elle établit
de nombreuses liaisons covalentes avec les autres composés. Les cellules ayant synthétisé de
la lignine meurent. La vacuole est le plus grand compartiment de la cellule végétale, il
représente jusqu’à 80 % de son volume. Il est limité par une membrane, le tonoplaste, qui
présente de très nombreuses protéines responsables de différentes formes de transport. La
vacuole participe au maintien de la forme de la cellule et à sa croissance en exerçant une
pression de turgescence sur la paroi. Elle fonctionne comme un lysosome de cellule
animale en détruisant les organites usagés et certaines macromolécules, elle stocke les
déchets, mais aussi certains métabolites. En contrôlant les échanges d’eau avec le milieu
extérieur, la vacuole maintient la pression de turgescence et dans une moindre mesure le pH.
En occupant l’essentiel du volume cytoplasmique, la vacuole déporte latéralement tous les
organites (noyau, mitochondries, chloroplastes,…). Les chloroplastes sont soumis à un
mouvement de cyclose et se déplacent autour de la vacuole ce qui les expose successivement
à la lumière et améliorent la conversion énergétique.

3
Les chloroplastes sont des organites à double membrane à l’intérieur desquels se déroulent
la photosynthèse. Les thylacoïdes sont le siège des conversions énergétiques et le stroma le
lieu de fixation et de réduction du CO2.Une partie du glucose ainsi produit est oxydée dans
le hyaloplasme et les mitochondries pour alimenter la cellule en ATP, qui ne peut être stocké,
l’autre partie est utilisée comme précurseurs des autres synthèses organiques.
La compartimentation cellulaire a permis aux cellules animales et végétales eucaryotes
d’atteindre un volume très nettement supérieur à celui des procaryotes (1 000 à 10 000 fois).
Une bactérie mesure en moyenne 1-2 mm de longueur contre 20-50 mm pour une cellule
animale et 100-250 mm pour une cellule végétale. Cette compartimentation délimite des petits
volumes cellulaires dont la composition chimique, le contenu enzymatique, le pH… diffèrent
d’un compartiment à l’autre. De plus, les molécules s’y rencontrent rapidement et les
différentes réactions sont ainsi favorisées. Les membranes qui délimitent les organites
possèdent des différences dans leurs compositions lipidiques et protéiques qui leur
confèrent des rôles totalement différents.
Les chloroplastes et les mitochondries sont des organites semi-autonomes qui possèdent de
l’ADN et des ribosomes. On pense qu’ils proviennent de l’endosymbiose de procaryotes
primitifs qui ont transféré une partie de leur information génétique dans le noyau, en ont
perdu une autre et en ont conservé qu’une petite fraction.
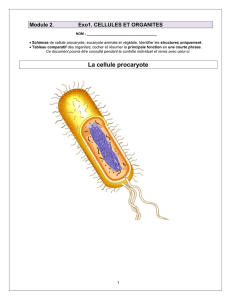
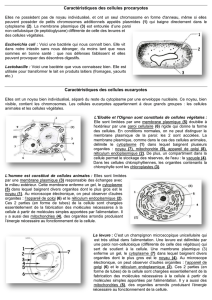
La cellule procaryote est la plus petite unité vivante, elle est capable d’assimilation, de
croissance et de reproduction. À l’échelle des temps géologiques, elle évolue. Une cellule
est un système thermodynamique ouvert qui, en utilisant l’information génétique qu’elle
possède, prélève matière et énergie dans l’environnement pour en faire sa propre substance
ordonnée assurant ainsi sa croissance et sa reproduction. Ce faisant, en dégageant de la
chaleur et des déchets, elle augmente considérablement l’entropie du milieu.
Chapitre 3 : Techniques d’études des cellules
La microscopie permet l’observation des structures des tissus et des cellules, elle ne
renseigne que très peu sur le fonctionnement cellulaire.
La microscopie photonique possède un grossissement maximal de 2 000 mais en pratique, il
n’excède pas 1 400 pour conserver des images nettes. L’observation de tissus ou de cellules
nécessite la réalisation de coupes minces de 2 à 10 µm que l’on colore de manière spécifique
afin d’augmenter les contrastes des différentes structures. La coloration de Feulgen colore
l’ADN en vert et les ARN en rouge. La coloration au camin-vert d’iode colore les parois
cellulosiques en rose et les parois lignifiées en vert.
Le microscope optique (MO) permet l’observation de cellules vivantes et le suivi de
molécules fluorescentes. Associé à la vidéomicroscopie, il permet de voir les mouvements
intracellulaires. Le microscope optique est aussi utilisé pour le suivi des molécules
radioactives.

4
Le microscope électronique (ME) atteint des grossissements conduisant à un pouvoir
séparateur de 0,2 nm soit une résolution 1 000 fois supérieure à celle du microscope optique.
Le pouvoir de résolution du ME permet l’observation des grosses protéines. L’observation
des échantillons nécessite des coupes ultrafines de quelques dizaines de nanomètres
d’épaisseur. Le contraste des préparations est renforcé par des imprégnations par des métaux
lourds opaques aux électrons. Les observations au ME se déroulent sous vide et
l’observation des cellules vivantes est impossible. Toutes les images sont en noir et blanc,
elles sont maintenant colorisées par des traitements informatiques. Le microscope
électronique à transmission (MET) produit des images à deux dimensions alors que le
microscope électronique à balayage (MEB) donne des images tridimensionnelles de plus
faible grossissement.
Les techniques de centrifugation trient les différentes fractions cellulaires en fonction de
leur densité. Ainsi, on isole les différents organites soit pour les observer au ME, soit pour
étudier leur biochimie structurale et fonctionnelle.
Les isotopes radioactifs (tritium) permettent de suivre les déplacements d’une molécule dans
une cellule. On utilise la technique du pulse-chasse. Durant le pulse de courte durée le
système biologique est approvisionné en éléments radioactifs, puis pendant la chasse, on
cherche la localisation de la radioactivité. La radioactivité est révélée par l’impression d’une
émulsion photosensible associée au dispositif optique.
Les anticorps monoclonaux fabriqués à partir des hybridomes sont des molécules qui
localisent de manière spécifique des protéines dans la cellule ou tout autre antigène. Ces
molécules sont ensuite marquées par un fluorochrome. Illuminé par un laser, les
fluorochromes émettent une lumière qui indique précisément les localisations de l’antigène
recherché.
La cytométrie de flux est une technique de tri des cellules dans une culture en fonction de
certaines de leurs caractéristiques révélées par le nombre de fluorochromes qu’elles ont
fixés.
Chapitre 4 : Etudes des micro-organismes
Les micro-organismes constituent un groupe totalement hétérogène d’êtres vivants qui sont
d’une taille inférieure au millimètre. La microbiologie est l’étude des micro-organismes,
cette discipline se caractérise par les techniques utilisées notamment pour ce qui concerne
leur culture et leur séparation.
Les micro-organismes comprennent tous les procaryotes (eubactéries et archées), les
levures (mycètes) et tous les protistes c’est-à-dire au sens restreint, tous les unicellulaires
eucaryotes autotrophes ou hétérotrophes au carbone. Les virus, bien que non vivants, sont
aussi un sujet d’étude des microbiologistes.
La culture des micro-organismes se réalise sur support solide (boîte de Pétri) ou en phase
liquide (flacon). Elle nécessite des conditions de stérilisation draconienne afin d’éviter des
contaminations. Les micro-organismes sont cultivés sur différents milieux qui permettent de
les isoler en fonction de leurs besoins nutritifs. Les milieux solides sont ensemencés à partir
d’un milieu liquide avec un étaloir ou râteau destiné à les répartir de manière uniforme sur
toute la surface de la gélose de la boîte. Chaque colonie se développant sur le milieu provient
d’un seul micro-organisme. Le dénombrement des colonies sur boîte est une méthode pour
compter les bactéries dans une culture.
En milieu liquide, la croissance d’une colonie d’unicellulaires se caractérise par une phase
de latence, une phase exponentielle, une phase stationnaire et une phase de déclin.

5
Le comptage des micro-organismes dans une culture se réalise selon plusieurs modalités. Les
méthodes directes dénombrent les cellules présentes dans le milieu alors que les méthodes
indirectes évaluent les modifications des caractéristiques du milieu durant la culture. Les
méthodes de dilutions successives sont utilisées soit pour ensemencer directement des boîtes
de Pétri, soit pour réaliser des dénombrements dans des cellules de comptage.
Les levures sont des mycètes unicellulaires qui se reproduisent essentiellement par
bourgeonnement ou division. Une de leur originalité est de se développer en conditions
aérobies ou anaérobies. En l’absence de dioxygène, elles fermentent et dégagent du
dioxyde de carbone et produisent de l’éthanol, dans ces conditions la croissance de la culture
est moins rapide.
Les protistes constituent un assemblage hétérogène de plusieurs lignées non directement
apparentées. Les coccolithophoridés constituent la craie déposée au crétacé, les diatomées et
les radiolaires sont pourvus d’un test siliceux et sont à l’origine des roches sédimentaires
formées sur les fonds océaniques, les foraminifères constituent d’excellents fossiles
stratigraphiques. D’autres, comme les paramécies, sont dépourvus de squelette, mais
possèdent une organisation très complexe. Certains protistes comme les euglènes passent
selon les conditions du milieu d’une autotrophie à une hétérotrophie au carbone. C’est dans
les protistes du type chlamydomonas que s’enracine la lignée verte à l’origine de tous les
végétaux terrestres.
Les bactéries sont des cellules procaryotes qui présentent une très grande diversité. Sous
leur apparente simplicité se cachent deux grands ensembles : les eubactéries et les archées.
Les eubactéries sont les bactéries communes et les archées sont des bactéries présentant de
très nombreuses particularités (structure et composition des membranes, organisation de
l’information génétique) qui de plus vivent dans les milieux les plus hostiles de la planète.
Les eubactéries se différencient sur la base de la coloration Gram en bactéries Gram + qui
présentent au-dessus de la membrane plasmique une importante paroi de peptidoglycanes et
en bactéries Gram − qui possèdent, en plus de la membrane plasmique, une bicouche
lipidique supplémentaire au-dessus d’une fine couche de peptidoglycanes.
1
/
5
100%