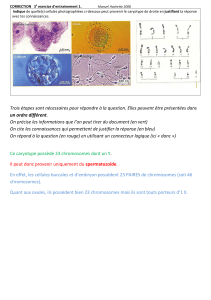
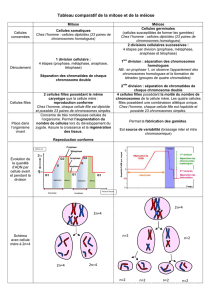
La Division cellulaire

La Division cellulaire
Possibilité à partir d’une cellule d’en obtenir, dans le cas général, deux identiques à la cellule
de départ. Possible pour toutes les cellules somatiques
Sauf : (neurones, musculaire striées, photorécepteurs de l’œil…)
Chez les unicellulaires : plus il y a à manger, plus on se divise rapidement. Chez les animaux,
il existe des mécanismes de contrôle assez complexes. Il y a nécessité de facteurs de
croissance.
Possibilité de division limitée dans le temps : une centaine de Génération après elle perd ses
capacités de contrôle. Excepté pour les cellules cancéreuses.
Pour un adulte, on observe une homéostasie cellulaire, à savoir un renouvellement équilibré
des cellules assurant l’intégrité de l’individu.
Cette capacité de se diviser, la mitose, n’est qu’une étape particulière de la vie des cellules, du
cycle Cellulaire.
I. Le Cycle Cellulaire (p36)
Deux grandes parties dans la vie d’une cellule :
- Entre deux divisions = Interphase
- Phase de division = Mitose
Plus grande partie de la vie d’une cellule (90%) : Interphase
La succession de ces deux grandes parties = cycle cellulaire
Interphase = 90% du cycle
- Période de croissance de la cellule : synthèse de toutes les molécules organiques
nécessaires (protéine, ARN, ADN, Lipides, Glucides, …)
- La Cellule grossit jusqu’à un volume double, donnant naissance à deux cellules au
volume égal à celui de la cellule mère.
Toutes les molécules sont synthétisées tout au long de l’interphase sauf pour l’ADN :
seulement pendant la période S (elle atteint strictement le double)
Généralement, la quasi-totalité des cellules eucaryotes possèdent 4 phases : tétraphasique.
G1 - S - G2 - M
On trouve un cycle biphasique au cours des premières divisions ȻR du zygote, par exemple.
Il existe également des cellules « hors cycle », certaines sont partie en phase G1
Sous une stimulation, elles peuvent revenir : G0-1, d’autre ne pourront pas : G0-D
A. La Phase G1
La phase la plus longue du Cycle, dont la durée est la plus variable.
C’est donc sur cette durée que la distinction cycle court/cycle long sera faite (pour un même
organisme).
C’est la phase de synthèse de tout un tas de produits, comme les produits de sécrétion.
C’est une période absolument essentielle pour le déroulement de la division qui va suivre. Elle
comporte un point particulier de contrôle : le point de restriction.

→ Lorsque la cellule dépasse ce point, alors elle va se diviser : phénomènes de
phosphorylation de protéines : activité d’enzymes de phosphorylation : kinases (régulée par
des protéines : cyclines).
Il existe quelques cas particuliers où l’on n’a pas 4 phases. N’existe par exemple que synthèse
et mitose c’est donc des cycles biphasiques. On les retrouve à propos des toutes premières
divisions qui succèdent à la fécondation.
B. La Phase S
Synthèse de l’ADN
4 copies identiques de chaque chromosome une fois la duplication effectuée.
Temps peu variable, 6-8h.
La réplication se fait à partir de points d’initiation. Ces réplications ne sont pas tout à fait
synchrones à partir de chaque point d’initiation. Le tout se chevauchera à la fin de manière à
avoir un ADN complet.
Cela donne lieu à des réplicons qui se chevaucheront pour achever la réplication.
Nombre moyen de réplicons : environ une centaine de réplicons par chromosome.
Moyenne de 50pb.s-1
Chez un modèle de cellule eucaryote unicellulaire : levures (Saccharomyces Cerevisire). Il
permet d’avoir des idées sur l’endroit où se placent ces points d’initiation.
Distribution aléatoire ou pré-établie ?
Séquence ARS (réplication autonome), on démontre que la position de ces séquences n’est
pas aléatoire, on en conclut donc que les points d’initiation pour toutes les cellules eucaryotes
ne sont pas positionnés aléatoirement.
On retrouve des séquences de consensus d’environ 11pb très concentrée en A-T.
Chez la levure il existe plus de séquence ARS que nécessaire pour répliquer l’ADN.
Il y a plus de points d’initiation possibles : certains sont sûrement muets.
Ces points d’initiation ne sont pas répartis de manière homogène le long du chromosome.
Il existe des foyers/usines de réplication où vont se trouver de manière très importante ces
points d’initiation. Ces foyers sont probablement associés à des éléments de la matrice
nucléaire, élément qui coordonne l’activité des différents points d’initiation
Il y a un schéma de réplication : au début on débute par l’euchromatine, puis la chromatine
associée au nucléole puis l’hétérochromatine. A chaque cycle, on re trouve la même séquence.
Au niveau d’un chromosome donné, visible, on peut définir des zones de réplication précoces
(télomères, zones riches ne gènes [bandes R]), réplication plus tardives pour les zones pauvres
en gènes (Bandes G), puis enfin, les zones centromériques.
D’un point de vue moléculaire, la réplication est semi-conservative, on a un brin ancien et un
néoformé dans les deux cellules filles.
Initiation de la duplication de l’ADN se fait donc en différents points de types ARS au niveau
desquels il existe tout ce qui est nécessaire à la duplication de l’ADN. Complexe
multienzymatique : réplisome (ensemble de ces enzymes).
Réplication qui intéresse tout d’abord l’euchromatine (apparait décondensée), puis la

chromatine associée au nucléole (autour des nucléoles) et les régions hétérochromatiques.
Au niveau de chaque chromosome, il existe des zones de réplications précoces : télomères et
zones riches en gènes (bandes R), des zones plus tardives (zones pauvres en gènes), avec en
dernier la zone centromérique.
Duplication selon un mode semi-conservatif (1brin parental et 1 néoformé).
Duplication de l’ADN, mais également des protéines qui l’accompagnent : histone ou autre.
Non-histones : pas de schéma très net. Elles s’associent à l’ADN et aux protéines histones.
Histones : genèse des nouveaux nucléosomes se fait via un mode partiellement conservatif.
Néonucléosomes se mettent en place au niveau de la fourche de duplication :
dissociation partielle des nucléosomes qui existe au niveai de cette zone : (H2A,
H2B, H3 , H4). Le comportement de ces histones n’est pas identique.
- H3 et H4 restent liés à l’ADN et seront distribués de manières aléatoires entre les deux
molécules d’ADN néo-formés : mode conservatif.
Nouveau nucléosomes : 50/50 de nouveau ou ancien H3 – H4
- Les nouveaux nucléosomes ne comportent quasiment que des H2A et H2B néoformés.
Il existe des quantités d’histones très importantes qui sont fabriquées pendant la phase S.
90% de la transcription des gènes des histones s’effectuent durant la phase S.
10% : ARNm des histones qui peuvent être fabriqués en dehors de la phase S, et ont une durée
de vie relativement longue.
C. La Phase G2
Phase post synthétique, qui précède la mitose.
Durée relativement variable : 4-5H
Ce stade nécessite un certain nombre de synthèses protéiques.
Si l’on bloque de manière artificielle la synthèse de protéine via des antibiotiques ou autre, la
future division ne pourra se produire.
Les protéines synthétisées pendant cette phase sont impliquées dans la condensation de la
chromatine.
Ces protéines sont des protéines appelées SMC, permettant la mise en place des microtubules
autour des couples centriolaires.
Evénement morphogénétiques et biochimiques permettant donc la division.
II. La Division Cellulaire : Phase M
Elle implique une disparition de la membrane nucléaire.
Ce sont des mitoses dites « ouverte ».
On appelle parfois cette situation, une division cellulaire de type Orthomitose (classique).
Certaines cellules se divisent en conservant leur enveloppe pleuromitose (cachée)
Cellules animales des mitoses de type Astérienne
Cellules végétale : anastrale
- Division du Noyau : Caryocinèse
Evénements morphologiquement visibles autour du noyau.
- Division du Cytosol : Cytodiérèse / Cytocinèse

A. La Mitose
1) La Prophase
15-20min
Enveloppe du noyau est toujours visible, on peut voir les chromosomes (premier signe
d’entrée en division). On peut donc les compter : 2n chromosomes.
Si on regardait ces chromosomes en prophase, on verrait qu’ils sont constitués de deux
chromatides : très peu visibles en MO. Elles sont le résultat de la duplication.
Elles sont séparées sauf au niveau de leur centromère.
Différenciation d’une structure morphologiquement visible : deux kinétochore
diamétralement opposés correspondant à une association avec les chromatides.
On peut les mettre en évidence via des Ac pendant l’interphase, mais ils sont visibles qu’au
début de la Mitose.
Modèle cellulaire : levure (saccharomyciès …. CF plus haut)
La condensation chromatique est due à l’action d’un complexe protéique qui est constitué de
5 peptides : le complexe condensine.
C’est lui qui explique la condensation des chromosomes.
Parmi ces 5, il existe deux peptides de type SMC qui vont jouer un rôle au tout début de la
division cellulaire, qui Maintienne la Structure des Chromosomes, avec une activité
ATPasique. Smc 2 et Smc 4 chez la levure.
Ces deux molécules sont associées en dimères.
Les 3 autres protéines sont des sous unités auxiliaires, qui ne sont pas de type SMC. Ce sont
des CAP, nécessaire à l’activité ATPasique des SMC.
Le complexe condensine est lié à la chromatine uniquement pendant la mitose, mais elle ne
s’y associe qu’au tout début de la division cellulaire.
Hydrolyse de l’ATP va permettre un surenroulement de la fibre nucléosomique, dont la
mécanique reste hypothétique.
Agencement de la fibre nucléosomique enroulée se surenroule.
Rapprochement des nucléosomes voisins, conséquence de la phosphorylation des Histones H1
Phosphorylation d’une autre espèce d’histone : H3, au début du cycle cellulaire.
Tout ceci nécessite un réarrangement de la fibre nucléosomique.
Topoisomérase II participe à cette condensation pour rendre visible les chromosomes en MO.
2) La Pré- Prométaphase = Fin Prophase
Milieu-Fin de prophase étant marqué par une interruption du processus d’endo- et exocytose.
Ne dure que quelques minutes.
Événement essentiel : rupture de l’enveloppe nucléaire conséquence de la phosphorylation des
lamines de la lamina provoquant la désorganisation de la lamina et donc de l’enveloppe
nucléaire, les filaments de l’enveloppe devenant indépendants les uns des autres. Les
constituants de cette enveloppe sont donc libres.
Lamines Type B reste attaché au système membranaire réticulaire
Lamines A et C deviennent libres.
Il n’y a donc plus de noyau au sens strict du terme.
Pendant ce stade, on va voir se différencier de manière bcp plus nette les kinétochore de

chaque chromosome.
État ultime de condensation des chromosomes.
Kinétochores = Aspect tripartite. Deux couches opaques aux électrons séparés par une couche
claire.
Plaque interne (chromatine) et externe (microtubules kinétochoriens = fibres).
Seulement certains nucléosomes d’une chromatine donné peuvent permettre l’attachement à
une plaque interne. Ils possèdent une histone H3 un peu différente.
15aine de protéines qui appartiennent à cette plaque interne.
Au niveau de la plaque externe, il existe là aussi des protéines de type motrices associées aux
microtubules (MAPs) : dynéines et des KRP (type kinésine).
Rôle des protéines apparentés, de type kinésine, mais jamais trouvée dans des protéines
impliquées dans le transport de vésicule !
Naissance de microtubule nés de couples centriolaires : kinétochore : microtbules
kinétochoriens « capturés » par les kinétochores. Extrémités + collés sur la plaque externe. Il
existe environ 1 microtubule par kinétochore pour les levures.
Les fibres kinétochoriennes vont placer les chromosomes sur un seul plan de division.
Ces fibres vont, au niveau des deux kinétochore diamétralement opposés, faire des
phénomènes de traction inégaux sur les chromosomes : plus les fibres sont longues, plus la
traction sera forte vers le centre nucléolaire qui leur a donné naissance.
Ils sont là pour positionner les les chromosomes sur un même plan.
Polymérisation / dépolymérisation des microtubules.
Tous ces déplacements sont asynchrones.
Cette prométaphase ne dure que quelques minutes.
3) La Métaphase
30-40min
Tous les chromosomes partiellement dédoublés alignés sur le plan équatorial.
Les kinétochores sont tournés de manière régulière vers les deux pole du fuseau de division.
Les chromosomes sont toujours en train de bouger, avec un équilibre statique même s’ils sont
sujet à des oscillations.
Concernant le fuseau de division, c’est le moment où l’on voit le plus de microtubules dont on
distingue 3 classes :
- Kinétochoriens (= 80%), qui ont la plus grande longueur
- Polaires, pas en relation avec les kinétochores, qui se chevauchent au pôle équatorial.
- Libres : perte de relation avec le centriole.
- Astériens : courts projetés vers mb plasmique. Rôle dans positionnement des
Chromosomes par rapport à la membrane plasmique.
 6
7
8
9
6
7
8
9
1
/
9
100%