Master de recherche des sciences Biologiques et médicales

17
Etude bibliographique
I. Eléments de biologie cutanée
Nous allons rappeler ici l’architecture générale de la peau avant de nous intéresser plus
particulièrement au système immunitaire cutané.
A. Structure de la peau chez les mammifères
La peau constitue une interface séparant le milieu extérieur du milieu interne de
l’organisme. Elle possède entre autres un rôle de protection mécanique et immunitaire.
La peau est constituée de trois couches : l’épiderme, le derme et l’hypoderme. (Figure 1)
Figure 1 : Schéma d’une peau saine (d’après Ibisch, 2002).
Couche cornée
Couche
granuleuse
Couche
épineuse
Couche basale
Membrane
basale
Derme
et
hypoderme
Cellule de
Langerhans
Kératinocyte
Cellule de
Merkel
Mélanocyte
Fibroblaste
Capillaire

18
L’épiderme est constitué majoritairement de kératinocytes, mais aussi de mélanocytes,
de cellules de Langerhans, et de cellules de Merkel. Les kératinocytes possèdent un rôle
structural : ils élaborent la kératine, lipophile, qui limite les pertes hydriques et qui est
responsable du maintien de l’épiderme. Par leur renouvellement constant et leurs jonctions
serrées, les kératinocytes sont aussi la première défense mécanique contre la pénétration
d’agents pathogènes. Ils exercent aussi un rôle actif lors d’inflammation cutanée puisqu’ils
peuvent synthétiser, sous l’action d’agents divers comme l’INFγ, différentes molécules
(Lloyd, 1990, Williams et Kupper, 1996) :
- des cytokines en quantité suffisante pour initier une inflammation cutanée
(IL-1 et TNFα)
- des cytokines de type Th1 et Th2 (IL-10 et IL-12 (seulement la p35)
- des facteurs de croissance pour les lymphocytes T (IL-7 et IL-15)
- les cytokines IL-8, TGFβ...
- des chimiokines
- des molécules du CMH II
- des molécules ICAM -1 permettant l’adhésion des lymphocytes T
Ainsi les kératinocytes peuvent se comporter comme cellules présentatrices
d’antigènes dans les inflammations cutanées : il s’agit du concept de « kératinocyte activé ».
Ils peuvent de cette façon s’ajouter aux cellules de Langerhans lors d’inflammation pour
présenter les antigènes.
Les cellules de Merkel sont des mécanorécepteurs à réponse lente, les médiateurs
qu’elles libèrent modulent l’activité des terminaisons nerveuses voisines.
Les mélanocytes, par la synthèse de mélanine, sont responsables de la teinte de la peau
et de la protection vis-à-vis des UV.
Le derme assure les apports en nutriments de l’épiderme grâce à sa vascularisation. Il
est essentiellement composé de fibroblastes qui élaborent une trame de fibres conjonctives
apportant la résistance et l’élasticité à la peau. Il comporte aussi les cellules de la lignée
granulocytaire et des cellules dendritiques.
A l’interface entre le derme et l’épiderme, on trouve la membrane basale composée en
majeure partie de collagène de type IV, qui permet l’adhésion derme-épiderme mais qui
intervient également dans la cicatrisation en donnant une trame de déplacement aux
kératinocytes basaux.

19
L’hypoderme est un tissu conjonctif plus lâche que le derme. Richement vascularisé, il
comporte des adipocytes et joue ainsi un rôle dans l’isolation thermique et le stockage des
réserves énergétiques de l’organisme.
La peau est, de plus, recouverte d’un mince film lipidique constitué de sébum élaboré
par les glandes sébacées et des sécrétions des glandes sudoripares. Le sébum, de composante
lipidique (acide linoléique), possède des propriétés bactériostatiques et permet de stabiliser le
pH cutané (Bouwstra et al. 2000).
La peau est donc un organe capable de limiter la prolifération bactérienne et les
pertes hydriques. Cependant, elle possède aussi tous les éléments nécessaires pour initier et
amplifier une véritable réponse immunitaire.
B. Système immunitaire cutané
En plus d’avoir un mode de défense passif par renouvellement constant, la peau
possède un système immunitaire propre qui permet l’initiation des réponses immunitaires.
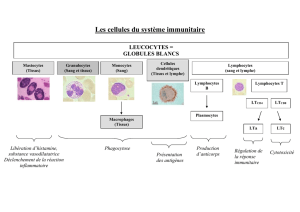
1) Les cellules présentatrices d’antigènes (CPA)
Les premières cellules capables de capter les antigènes et de les présenter aux
lymphocytes Th sont les cellules de Langerhans et les cellules épidermiques dendritiques
inflammatoires (IDEC). Les cellules de Langerhans sont reconnaissables en microscopie
électronique grâce aux granules de Birbeck (granules en forme de raquette). Ces cellules
migrent par voie lymphatique pour présenter l’antigène dans les ganglions. Les cellules de
Langerhans sont CD1a+/CD207+, les cellules épidermiques inflammatoires, elles, sont
caractérisées par la présence du récepteur au mannose (CD206+) et sont ainsi capables
d’endocytose de composants bactériens (Wollenberg et al. 1996).
Comme nous l’avons vu, les kératinocytes sont capables, eux aussi, d’exprimer le
complexe majeur d’histocompatibilité de classe II et ainsi de se comporter comme une
cellule présentatrice d’antigène dans un contexte inflammatoire.

20
2) Les lymphocytes
Les lymphocytes T sont issus du thymus, ils migrent, après sélection positive et
négative, dans les tissus périphériques. Au niveau cutané, on retrouve les lymphocytes
épithéliotropes, les lymphocytes T CD4+ (ou lymphocytes Th) qui reconnaissent le CMH de
classe II par l’intermédiaire de leur TCR. Une fois activés, ils orientent la réaction
immunitaire vers une immunité à médiation cellulaire ou humorale. (Figure 2)
Les lymphocytes B, originaires de la moelle osseuse, sont très peu présents dans le
derme non inflammatoire, mais se retrouvent au niveau des ganglions drainants.
3) Les granulocytes
Ils sont en nombre restreint dans une peau saine. Les granulocytes éosinophiles ont
pour rôle principal la lutte contre les parasites. Cependant ils sont capables, même si ce n’est
pas leur fonction la plus importante, d’endocytose comme les granulocytes neutrophiles (Roitt
et al. 1994). Ces derniers jouent un rôle non négligeable lors d’infection bactérienne et
constituent la majeure partie du pus.
Les granulocytes basophiles présentent des similitudes avec les mastocytes : si le
deuxième est tissulaire et se retrouve dans le derme, ils possèdent tous deux des récepteurs
appelés FcεR et contiennent les mêmes médiateurs dans leurs vésicules comme : l’histamine,
la sérotonine, l’héparine, des protéines neutres comme la tryptase et la chymase, et des
chondroïtines sulfates (Roitt et al. 1984).

21
Figure 2 : Le paradigme Th1 / Th2 (Prélaud, 1999).
Nous venons de voir que la peau de Mammifère possède de nombreux éléments lui
permettant de se défendre contre la pénétration d’allergènes cutanés. Cependant, dans le
cas de la dermatite atopique par exemple, ou encore des allergies de contact, on observe une
réponse immunitaire inappropriée suite à un contact avec un allergène déclenchant ainsi la
maladie. Il s’agit d’une des rares maladies (avec quelques autres maladies allergiques comme
l’asthme, la dermatite de contact et la rhinite allergique) dans lesquelles la pathogénie repose
sur la réaction immunitaire qui survient suite à la présence d’une protéine exogène à la surface
de la peau ou d’une muqueuse. Nous allons détailler dans cette deuxième partie, la
physiopathologie de la dermatite atopique.
Th0
IL-12
Th2 Th1
INF
γ
IL-2
IL-4
IL-10
IL-13
IL-4, IL-6
IL-4
IL-10
IL-13
LB
IgEIgG
Macrophage EosinophileMastocyte
CPA
IL-3
IL-4
IL-10
IL-5
IL-13
IL-4
CPA : Cellule présentatrice d’antigène Ig : Immunoglobuline
IL : Interleukine
LB : Lymphocyte B
Th : Lymphocyte T helper
Ce schéma présente de manière simplifiée le devenir possible des lymphocytes T helper, les
différentes cytokines sécrétées selon les cas et l’action des différents sous-types Th sur le
système immunitaire. Par souci de simplification, les boucles de régulation et l’action des CP
A
n’ont pas été incluses dans ce schéma.
1
/
5
100%