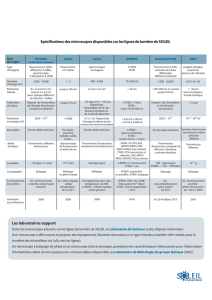
le photoeblouissement

LE PHOTOEBLOUISSEMENT
Philippe PAGO
Lorsqu'un fluorochrome est exposé à des rayons lumineux, la fluorescence qu'il émet diminue
progressivement dans le temps (photoextinction). Ce phénomène physique représente très
souvent une gène pour les analyses morphologiques et quantitatives en fluorescence, en
particulier pour celles faisant appel à des techniques de cytométrie (flux, image, confocal).
Le photoéblouissement est une utilisation volontaire de cette propriété: une source lumineuse
est utilisée pour éteindre l'émission de fluorescence d'un fluorochrome. Il est mis à profit dans
une application particulière de la cytométrie en image: l'étude de la récupération de fluorescence
après photoéblouissement (Fluorescence Recovery After Photobleaching: FRAP). Une telle
technique d'investigation permet d'étudier les déplacements moléculaires à l'échelon tissulaire,
cellulaire et infracelluaire .
PRINCIPE GENERAL
La molécule, dont on veut étudier les déplacements, doit être fluorescente ou rendue
fluorescente par couplage à un fluorochrome. Elle est incorporée dans la structure biologique
d'intérêt (par simple incubation, micromanipulation...).
Un faisceau laser puisé est utilisé pour éteindre la fluorescence dans une région bien déterminée
de la préparation.
La récupération de la fluorescence dans la région photoéblouie est ensuite étudiée en fonction
du temps. Cette récupération de fluorescence est liée au déplacement des molécules
fluorescentes depuis les zones non photoéblouies vers les zones photoéblouies.
MATERIEL
Bien que différentes techniques de cytométrie en image (microvidéomicroscopie en particulier)
puissent être utilisées pour étudier la récupération de fluorescence après photoéblouissement, le
cytomètre en image à balayage laser ACAS 570 de Méridien (Ronot et Raymond, 1991;
Métézeau et al., 1993) se révèle particulièrement performant dans cette application. La
préparation est placée sur la platine motorisée d'un microscope inversé. Le faisceau laser est à
énergie puisée (pour contrôler la puissance et la durée de l'excitation lumineuse) et à trajet fixe;
il éclaire, au travers de l'objectif du microscope, successivement chacun des points (champ,
cellule ou portion de cellule) de la préparation grâce au déplacement en X et en Y de la platine.
Les lumières de fluorescence (2 simultanément au maximum) émises en chacun des points de la
préparation sont transmises, au travers de l'objectif et d'un ban optique, à des
photomultiplicateurs qui les quantifient. L'équipement informatique du cytomètre établit des
images, en pseudo-couleurs, d'intensité de fluorescence point par point.
Pour l'étude de la récupération de fluorescence après photoéblouissement, la procédure est la
suivante. Une image d'intensité de fluorescence pré-photoéblouissement est acquise. Sur cette
image est délimitée la ou les zone(s) à photoéblouir. Le photoéblouissement est réalisé sous
contrôle informatique et une image d'intensité de fluorescence post-photoéblouissement est
immédiatement acquise. Par balayages successifs de la préparation, des images d'intensité de
fluorescence sont ensuite acquises à intervalles de temps réguliers. En fin d'expérimentation,
l'ordinateur affiche des images d'intensité de fluorescence et un diagramme représentant
l'évolution de l'intensité de fluorescence en fonction du temps, ceci dans différentes zones
d'intérêt définies par l'opérateur (figures 1 à 3).

Les différents de l'expérimentation (nombre et puissance des spots de photoéblouissement,
nombre de points de mesure en X et en Y, distance entre 2 points de mesure, vitesse de
balayage, intervalle entre 2 balayages et nombre total de balayages post-photoéblouissement...)
sont définis en fonction de la cinétique du déplacement moléculaire à étudier.
PRINCIPALES APPLICATIONS
Les principales applications de l'étude de la récupération de la fluorescence après
photoéblouissement sont illustrées à partir d'exemples issus d'une revue de la littérature de ces
deux dernières années.
1) Diffusion de molécules dans les tissus
Haller et Saltzman (1998) ont utilisé la FRAP pour étudier la diffusions de facteurs de
croissance à visée thérapeutique dans le système nerveux central.
2) Diffusion de molécules dans le noyau des cellules
La FRAP a été utilisée pour étudier les mouvements intranucléaires d'oligonucléotides utilisés
en cytogénétique moléculaire (Politz et al., 1998) et la dynamique de la chromatine dans le
noyau interphasique (Abney et al., 1997).
3) Diffusion de molécules dans le cytoplasme des cellules
3 - 1: Viscosité cytoplasmique
En ayant recours à la FRAP et à des macromolécules fluorescentes calibrées (Seksek et al.,
1997) ou à la GFP (Swaminathan et al., 1997), et à partir de l'analyse de la diffusion de ces
sondes, la viscosité du cytoplasme, dans différentes conditions, a pu être étudiée.
3 - 2: Transport intracellulaire de protéines. de lipides ou de vésicules
La FRAP a été utilisée pour étudier i) la mobilité intracellulaire de la tyrosine kinase TrkA et de
facteurs de régulation (Wolf et al., 1998), ii) le transport intracellulaire de la transferrine (Azizi
et Wahl, 1997; Wahl et Azizi, 1997), iii) le rôle de la différenciation cellulaire et de la protéine
intestinale liant les acides gras dans la capture et la diffusion intracellulaire des acides gras
(Atshaves et al., 1998) et iv) la mobilité des vésicules synaptiques dans les terminaisons
nerveuses (Kraszewski et al., 1996).
4) Diffusion de molécules dans les membranes des cellules
4 - 1: Fluidité membranaire et diffusion latérale de lipides dans la membrane plasmique
De nombreux travaux illustrent cette application de la FRAP: étude de l'influence sur la fluidité
de la membrane plasmique des lipides diététiques (Clamp et al., 1997; Tappia et al., 1997);
étude des effets de l'éthanol et du ganglioside GM1 sur la fluidité membranaire des cellules de
la crête neurale (Chen et al., 1996); étude des effets d'une infection par les prions sur la fluidité
membranaire des neurones (Wong et al., 1996); étude in vitro des modifications de la
diffusibilité des lipides dans la membrane du spermatozoïde lors de la capacitation (Smith et al.,
1998); étude de la mobilité latérale des lipides de la membrane plasmique des spermatozoïdes:
hétérogénéité entre domaines de surface et rigidification après mort cellulaire (Ladha et al.,
1997); étude des effets des acides gras sur la diffusion latérale des lipides dans la membrane
plasmique des kératinocytes (Fulbright et al., 1997); étude de la distribution et des propriétés
biophysiques de différents lipides à la surface de parasites (Redman et Kusel, 1996).
4 - 2: Diffusion latérale de protéines dans la membrane plasmique
Dans ce contexte, ont été étudiés i) la mobilité latérale d'oligomères band 3 dans la membrane

plasmique d'érythroblastes (Hanspal et al., 1998), ii) la diffusion latérale du récepteur Con-A
dans la membrane plasmique de cellules musculaires striées (Zs-Nagy et al., 1998), iii) la
détection de l'immobilisation de ligands due à leur liaison à des récepteurs et son application à
la signalisation cellulaire, à la thérapie génique et aux drogues ciblées (Berk et al., 1997), iv) la
mobilité latérale d'intégrines dans la membrane plasmique des mégacaryocytes (Schootemeijer
et al., 1997), v) la diffusion latérale au niveau de la membrane plasmique de molécules du
complexe majeur d'histocompatibilité de classe 11 et les actions sur cette diffusion d'agonistes
et de ligands altérés (Qiu et al., 1996) et vi) les effets de différents peptides du métabolisme
oxydatif sur la mobilité latérale du récepteur au complément de type I au niveau de la
membrane plasmique du neutrophile (Rasmusson et al., 1996).
4 - 3: Autres membranes cellulaires
La FRAP a été utilisée pour mettre en évidence une solution de continuité transitoire au niveau
de la membrane du réticulum endoplasmique au cours de la fécondation (Terasaki et al., 1996)
et étudier la mobilité diffusionnelle des protéines membranaires du Goigi (Cole et al., 1996).
5) Communications gap jonctionnelles
La FRAP a été également très utilisée pour rechercher et surtout étudier le rôle de différents
facteurs sur les communications intercellulaires gap jonctionnelles. Quelques exemples de
travaux illustrent cette application: implication transitoire des communications gap
jonctionnelles dans la fusion des myoblastes de rat nouveau né (Constantin et al., 997); étude de
la régulation des communications gap fonctionnelles durant la différenciation du trophoblaste
humain (Cronier et al., 1997); étude des effets de la concentration en glucose et de la
phosphorylation des protéines sur les communications intercellulaires des cellules pigmentaires
de la rétine en culture (Stalmans et Himpens, 1997); inhibition réversible par le tamoxifen des
communications gap fonctionnelles de myocytes cardiaques en culture (Verrecchia et Herve,
1997); étude des effets d'hormones lutéotrophiques et lutéolytiques sur les communications
intercellulaires gap fonctionnelles entre cellules lutéales à différents stades du développement
lutéal (Grazul Bilska et al., 1996); recherche de communications gap fonctionnelles entre
cellules de la granulosa et de facteurs les inhibant ou les activant (Furger et al., 1996); étude des
effets de différentes substances sur les communications gap fonctionnelles entre cellules
myométrales humaines immortalisées (Burghardt et al., 1996).
6) Autres applications en biologie
De nombreux travaux (non cités ici) font appel à la FRAP pour étudier la mobilité moléculaire
dans des modèles artificiels ou naturels de films biologiques mono-ou-bi-couches.
UN EXEMPLE D'APPLICATION DE LA FRAP: RECHERCHE DE
COMMUNICATIONS INTERCELLULAIRES ENTRE CELLULES SENSORIELLES
IMMATURES DE LA COCHLEE; ETUDE CRITIQUE
Cette étude a été menée en collaboration avec Marc Lenoir et Chantal Ripoll, à l'unité
INSERM 254 et au Centre Régional d’imagerie Cellulaire de Montpellier.
Introduction
Les cellules ciliées externes (CCE) de la cochlée sont des structures sensori-motrices douées de
propriétés contractiles. Elles jouent un rôle clé dans la fonction auditive en amplifiant et en
filtrant la vibration sonore. Ces propriétés sont acquises à un stade du développement où
l'organe de Corti est encore immature et les CCE sont jointives.
L'objectif de cette étude était de mettre en évidence l'existence éventuelle de communications
entre CCE immatures, les communications intercellulaires contrôlant la différenciation de
nombreux types cellulaires. Pour ce faire, nous avons appliqué, à des CCE immatures

maintenues en survie in vitro, la technique de récupération de fluorescence après
photoéblouissement, cette approche cytométrique permettant une exploration fonctionnelle des
communications intercellulaires. Nous avons abordé la question d'une façon critique en évaluant
notamment les effets, à l'échelon cellulaire, d'éventuelles lésions que pourrait engendrer le
faisceau laser.
Matériels et méthodes
Dix rats Wistar, âgés de 5 jours, ont été utilisés pour cette étude.
Après dissection des cochlées, les CCE étaient dispersées mécaniquement à l'aide d'une
micropipette par aspiration/rinçage de l'organe de Corti contenu dans une goutte de tampon
additionnée de thrombine. Chez le jeune rat, cette technique de dissociation en présence de
thrombine permet de recueillir des chapelets de CCE pouvant contenir jusqu'à 10 cellules.
Après sédimentation des cellules, une goutte de plasma de poulet était délicatement déposée
dans le milieu. La présence de thrombine dans ce milieu faisait coaguler le plasma et les cellules
étaient ainsi immobilisées sur le support.
Les CCE étaient ensuite incubées pendant 5 min en présence de calcéine AM (CAM) et
d'éthidium homodimère (EthD). La C-AM, sonde qui traverse les membranes cellulaires, est
estérifiée par les cellules vivantes leur conférant une fluorescence verte au niveau du
cytoplasme. L'EthD, sonde qui ne pénètre que dans les cellules aux membranes perméabilisées
(cellules mortes ou fixées), s'intercale dans l'ADN conférant une fluorescence rouge au noyau.
Les préparations étaient ensuite analysées avec un cytomètre en image à balayage laser (ACAS
570, Méridian). Avant et après chaque expérimentation, la viabilité des CCE était évaluée par
une analyse en double fluorescence (520 nm pour la C-AM; 620 nm pour l'EthD). L'extinction
de fluorescence était ensuite réalisée par exposition d'une ou plusieurs cellules ù une dizaine
d'éclairs laser très focalisés et de faible intensité (<1 mW). La récupération de fluorescence était
ensuite étudiée par balayage de la préparation toutes les 1 à 2 min pendant 10 à 30 min.
Résultats et discussion
1 ) Première condition expérimentale (figure 1 )
Après 5 min d'incubation en présence des sondes fluorescentes (C-AM et EthD), le milieu était
remplacé par du milieu frais. Aucune récupération de fluorescence après photo éblouissement
n'a été observée: les CCE adjacentes n'avaient donc pas échangé de C-AM avec les CCE a
éteintes ”. Il n'existe donc pas de communications gap jonctionnelles entre CCE immatures.
2) Deuxième condition expérimentale (figure 2)
Les CCE étaient maintenues dans le milieu contenant les sondes fluorescentes (CAM et EthD).
Une récupération de fluorescence était observée dans les 10 min suivant le photoéblouissement:
les CCE “ éteintes ” restaient donc capables d'estérifier la C-AM. Le faisceau laser, à la
puissance utilisée (< 1mW), ne génère pas de lésions vitales au niveau des CCE.
3) Troisième condition expérimentale (figure 3!
Une lésion était créée au niveau du pale basal de la CCE par un éclair laser très focalisé et de
très forte intensité (= 100 mW). Aucune extinction de fluorescence n'était observée initialement.
Après 5 à 20 min, une perte soudaine (en moins d'1 min) d'émission de fluorescence était
souvent observée au niveau de la CCE lésée. Environ 45 min plus tard, un marquage du noyau à
l'EthD était observé.
Ces données suggèrent que, dans un premier temps, les CCE entrent dans un processus de mort
cellulaire actif (des modifications intracytoplasmiques de pH, de concentrations ioniques... font
que l'émission de fluorescence de la C-AM est inhibée). Dans un deuxième temps, les CCE
subissent un processus de nécrose classique (les membranes cellulaires deviennent perméables à
l'EthD) vraisemblablement en raison des conditions in vitro dans lesquelles elles se trouvent. La

lésion provoquée par le faisceau laser induit donc initialement un processus de dégénérescence
active (apoptose ?) qui avorte secondairement dans notre modèle in vitro.
REFERENCES
1. Abney et al., 1997, J Cell Biol, 137, 459-1468
2. Atsheves et al., 1998, Am J Physiol, 274, C633-C644
3. Azizi et Wahl, 1997, Biochim Biophys Acta, 1327, 75-88
4. Berk et al., 1997, Proc Natl Acad Sci USA, 94,1785-1790
5. Burghardt et al., 1996, Biol Reprod, 55, 433438
6. Chen et al., 1996, Alcohol, 13, 589-595
7. Clamp et al.,1997, Lipids, 32,179-184
8. Cole et al., 1996, Science, 273, 797-801
9. Constantin et al.,1997, C R Acad Sci III, 320, 35-40
10.Cronier et al.,1997, Microsc Res Tech, 38, 21-28
11.Fu!bright et al.,1997, Exp cell Res, 233,128-134
12.Furger et al., 1996, Mol Hum Reprod, 2, 541-548
13.Grazul Bilska et al.,1996, Prostaglandins, 52, 285-302
14.Haller et Saltzman, 1998, Pharm res,15, 377-385
15.Hanspal et al.,1998, Blood, 92, 329-338
16.Kraszewski et al., 1996, J Neurosci, 16, 5905-5913
17.Ladha et al.,1997, J Cell Sci,110,1041 -1050
18.Métézeau et al.,1993, Path Biol, 41, 276-280
19.Politz et al., 1998, Proc Natl Acad Sci USA, 95, 6043-6048
20.Qiu et al., 1996, Immunol Lett, 53,19-23
21.Rasmusson et al., 1996, Biosci Rep, 16, 391-404
22.Redman et Kusel, 1996, Parasitology, 113,137-143
23.Ronot et Raymond, 1991, European Microscopy and analysis, 17, 27-29
24.Schootemeijer et al., 1997, Thromb Haemost, 77,143-149
25.Seksek et al., 1997, J Cell Biol, 138,131-142
26.Smith et al., 1998, Mol Reprod Dev, 50, 86-92
27.Stalmans et Himpens, 1997, Invest Ophtalmol Vis Sci, 38,1598-1609
28.Swaminathan et al., 1997, Biophys J. 72,1900-1907
29.Tappia et al., 1997, Mol Cell Biochem, 166, 135-143
30.Terasaki et al., 1996, Dev Biol, 179, 320-328
31.Vago et al., 1996, Anal Cell Pathol, 10,170
32.Verrecchia et Herve, 1997, Pflugers Arch, 434,113-116
33.Wahl et Azizi,1997, Biochim Biophys Acta, 1327, 69-74
34.Wolf et al., 1998, Biochemistry, 37, 3178-3186
35.Wolf et al., 1998, Biochemistry, 37, 3178-3186
36.Wong et al., 1996, Neurology, 47, 741-750
37.Zs-Nagy et al., 1998, Mech Ageing Dev,101, 257-268
 6
7
6
7
1
/
7
100%