Table des matières

xiv
Table des matières
Les auteurs v
Introduction à la biologie : Note de l’éditeur vii
Avant-propos ix
Remerciements x
CHAPITRE 1 Une vue d’ensemble
de la réponse immunitaire
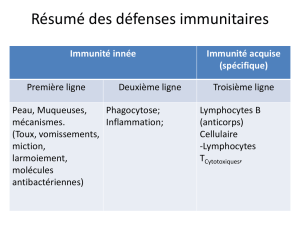
1-0 Les mécanismes de protection de l’immunité : un aperçu 2
Le système immunitaire nous protège contre les organismes infectieux
Les mécanismes immunitaires se répartissent en deux catégories :
l’immunité innée ou naturelle et l’immunité acquise ou adaptative
Définition opérationnelle d’un antigène
1-1 Les cellules du système immunitaire : différenciation dans la moelle osseuse 4
Les cellules du système immunitaire sont formées dans la moelle osseuse
Les cytokines produites par les cellules stromales de la moelle osseuse contrôlent
la différenciation des cellules immunitaires à partir de leur précurseur
hématopoïétique commun
Les réserves de cellules hématopoïétques de la moelle osseuse
sont mobilisées rapidement en réponse au stress
1-2 Les cellules du système immunitaire : caractéristiques fonctionnelles 6
La différenciation des cellules immunitaires peut comporter plusieurs phases
Les neutrophiles, les macrophages et les cellules dendritiques sont des cellules
phagocytaires
Les basophiles, les mastocytes et les éosinophiles libèrent des médiateurs
inflammatoires et cytotoxiques stockés dans des compartiments intracellulaires
Les lymphocytes détectent les antigènes au moyen de récepteurs variables
1-3 Les différents types de macrophages et de cellules dendritiques 8
Les macrophages et les cellules dendritiques sont les cellules
multifonctionnelles de l’immunité innée
Les macrophages se spécialisent différemment selon les tissus
Les cellules dendritiques ont des formes et des fonctions très variées
1-4 Sélection clonale de lymphocytes spécifiques d’antigène 10
L’adaptabilité et la sélectivité de l’immunité acquise sont assurées par des processus
de sélection qui agissent sur un répertoire hautement variable de récepteurs
Les différents récepteurs d’antigène sont produits par des réarrangements
de l’ADN lymphocytaire pendant l’ontogenèse
La sélection par l’antigène conduit à la tolérance du soi ainsi qu’à la reconnaissance
spécifique de presque tout organisme ou particule d’origine étrangère
1-5 Les molécules du complexe majeur d’histocompatiblité et la détection
de l’infection 12
Les lymphocytes T reconnaissent des fragments d’antigène transportés vers
la surface des cellules par les molécules CMH
Les molécules CMH sont des indicateurs de surface des protéines présentes
dans les compartiments internes des cellules
Les marqueurs de surface des différentes classes de cellules T reflètent les processus
de reconnaissance différents des molécules CMH de classe I et de classe II
1-6 Le système lymphoïde et la circulation des lymphocytes 14
Il existe deux types de tissus lymphoïdes
Les tissus lymphoïdes secondaires assurent la rencontre des antigènes et
des lymphocytes
Les chimiokines dirigent la migration des lymphocytes et organisent
les microenvironnements dans lesquels ils sont activés
Les tissus lymphoïdes secondaires sont stabilisés et leur formation peut être induite
par les signaux des cellules immunitaires
1-7 Architecture des tissus lymphoïdes secondaires 16
Les cellules dendritiques et les cellules dendritiques folliculaires récoltent
les antigènes en vue de leur reconnaissance par les cellules T et les cellules B
La rate est un filtre essentiel pour éliminer les antigènes présents dans le sang
Les antigènes entrent dans les tissus lymphoïdes muqueux en passant
par des cellules spécialisées
DeFranco_liminaires.qxp 1/28/09 1:43 PM Page xiv

xv
1-8 Le déroulement d’une réponse immunitaire 18
L’immunité innée et des classes spécialisées de lymphocytes constituent
une première ligne de défense contre les micro-organismes envahisseurs
Les lymphocytes T activés par les cellules dendritiques se différencient et migrent
vers les follicules de cellules B et les sites d’infection
Les lymphocytes B activés se différencient en plasmocytes et sécrètent
des anticorps
À la fin d’une réponse immunitaire, la vaste majorité des cellules spécifiques
d’antigène meurent et les cellules survivantes deviennent des cellules mémoire
quiescentes
CHAPITRE 2 Les molécules de signalisation et d’adhérence
du système immunitaire
2-0 Les molécules de surface et de signalisation des cellules immunitaires :
un aperçu 22
La migration et la communication des cellules immunitaires sont contrôlées
par des molécules spécialisées de la surface cellulaire
et par des molécules de signalisation
Les molécules d’adhérence jouent un rôle important dans la migration cellulaire
et dans les interactions entre cellules
De grandes familles de molécules médiatrices de signal contrôlent pratiquement
tous les aspects du développement et du fonctionnement du système immunitaire
2-1 La superfamille des immunoglobulines : aspects structuraux 24
Le domaine de type Ig est un élément structural important
de nombreuses protéines du système immunitaire
La superfamille Ig contient une variété d’éléments de reconnaissance moléculaire
du système immunitaire
2-2 Signalisation par les récepteurs de la superfamille des immunoglobulines 26
Les membres de la superfamille Ig sont des médiateurs de la reconnaissance
des antigènes et de nombreux effets fonctionnels des anticorps
Les récepteurs activateurs de la superfamille Ig transmettent le signal
par l’intermédiaire de tyrosine kinases intracellulaires
Les cellules immunitaires expriment des récepteurs inhibiteurs appartenant
à la superfamille Ig qui limitent les réponses immunitaires
2-3 Molécules d’adhérence de la superfamille des immunoglobulines 28
Les membres de la superfamille Ig jouent un rôle important
dans l’adhérence entre cellules immunitaires
La sous-famille CD2 participe à l’adhérence et à l’activation
des cellules du système immunitaire
Plusieurs membres de la superfamille Ig qui sont des ligands
des intégrines sont eux-mêmes attachés à des cellules
2-4 Le rôle des intégrines dans la fonction immunitaire 30
Les intégrines sont des médiateurs du contact entre cellules et entre cellules
et matrice extracellulaire
Les propriétés d’adhérence des intégrines sont modulées par d’autres signaux
qui parviennent à la cellule
L’engagement des intégrines régule la forme de la cellule et émet des signaux
qui supportent l’activation et la survie de la cellule
Les intégrines sont exprimées de façon différente selon la spécialisation
des cellules immunitaires
2-5 Les lectines de type C et la reconnaissance des hydrates de carbone 32
Les molécules qui reconnaissent les hydrates de carbone ont des rôles divers
dans le système immunitaire
2-6 Les cytokines et leurs récepteurs 34
Des médiateurs polypeptidiques sont l’outil principal de la régulation des réponses
immunitaires
Les cytokines agissent sur de nombreux types cellulaires de façon redondante
et coordonnée à la fois
Les cytokines peuvent être réparties en familles de molécules structuralement
apparentées qui utilisent des récepteurs homologues
2-7 Les récepteurs de cytokines qui transmettent le signal par la voie Jak-STAT 36
La famille des cytokines de type I est reconnue par une famille de récepteurs
conservés
Les interférons et l’IL-10 sont apparentés aux cytokines de type I et ils sont aussi
reconnus par des récepteurs qui transmettent le signal par la voie Jak-STAT
DeFranco_liminaires.qxp 1/28/09 1:43 PM Page xv

Les récepteurs de cytokines de type I et II transmettent le signal principalement
par la voie Jak-STAT
Les STAT déterminent largement la spécificité du signal des récepteurs de cytokines
Le signal Jak-STAT est régulé négativement par les protéines SOCS
2-8 Sous-unités communes dans les sous-familles de récepteurs de cytokines
de type I 38
De nombreuses cytokines et hématopoïétines sont reconnues par des récepteurs
apparentés structuralement qui ont des sous-unités communes
2-9 La superfamille TNF de régulateurs cellulaires 40
Les membres de la superfamille TNF régulent une variété de réponses immunitaires
et d’étapes du développement
Les membres de la superfamille des récepteurs de TNF transmettent
le signal par recrutements successifs
2-10 NF-κB et l’action inflammatoire des cytokines 42
NF-κB active l’expression des gènes de l’inflammation
NF-κB est régulé par des sous-unités inhibitrices
Certains hétérodimères de NF-κB s’auto-inactivent
2-11 Contrôle moléculaire de l’apoptose 44
L’apoptose a plusieurs rôles importants dans la fonction immunitaire
L’apoptose est déclenchée par une cascade protéolytique dans laquelle
interviennent les caspases
2-12 Régulation de l’apoptose par les membres de la famille Bcl-2 46
L’augmentation de la perméabilité de la membrane externe de la mitochondrie est
le principal déclencheur de la voie apoptotique intrinsèque
2-13 Les chimiokines et leurs récepteurs 48
Les chimiokines dirigent la migration des cellules du système immunitaire
2-14 Chimiokines, intégrines, sélectines et écotaxie (« homing »)
des cellules immunitaires 50
Les chimiokines collaborent avec d’autres molécules spécialisées pour guider
les lymphocytes et les leucocytes vers les tissus appropriés
Les sélectines sont des lectines de type C qui se lient à des structures glucidiques
présentes à la surface des cellules
Les lymphocytes circulants expriment un récepteur régulé qui dirige leur sortie
des tissus lymphoïdes
CHAPITRE 3 L’immunité innée
3-0 L’évolution et le fonctionnement de l’immunité innée : un aperçu 54
Les organismes pluricellulaires ont des mécanismes de reconnaissance immunitaire
innée conservés
L’immunité innée reste efficace malgré l’évolution rapide des microbes et des virus
Les mécanismes de reconnaissance de l’immunité innée sont antérieurs
à l’immunité acquise et ils lui donnent des instructions
3-1 Les barrières épithéliales contre l’infection 56
La couche épithéliale constitue une première barrière contre la plupart des infections
Les peptides antimicrobiens constituent un système de défense largement utilisé
3-2 La reconnaissance innée par les collectines et les ficolines solubles 58
Les collectines et les ficolines activent les mécanismes de protection immunitaire
innés à la surface des muqueuses et dans le sang
Les collectines et les ficolines ont une architecture commune mais des domaines
de reconnaissance différents
Deux surfactants pulmonaires sont des collectines
La lectine liant le mannose fournit une protection immunitaire innée contre certaines
infections bactériennes
3-3 Une vue d’ensemble du système du complément 60
La cascade du complément fait le lien entre la reconnaissance des microbes
par des facteurs solubles et les mécanismes qui les détruisent
Des déficits congénitaux du complément résultent en une tendance
au développement d’infections bactériennes sévères et à une susceptibilité
à des maladies immunes complexes
3-4 L’activation de la cascade du complément 62
Les étapes initiales de l’activation du complément par les voies classique
et des lectines ont beaucoup d’éléments communs
L’activation du complément est essentiellement circonscrite aux surfaces cellulaires
xvi
DeFranco_liminaires.qxp 1/28/09 1:43 PM Page xvi

La voie alternative d’activation du complément fournit une boucle d’amplification
qui est activée à la surface des cellules
3-5 Les actions effectrices et les récepteurs du complément 64
La cascade du complément stimule l’inflammation, la phagocytose, l’attaque
membranaire et la production d’anticorps
Des récepteurs du complément interviennent dans beaucoup
des effets du complément
3-6 Régulation du complément et stratégies d’évasion microbiennes 66
L’activation du complément est régulée par des facteurs solubles et des inhibiteurs
liés à la membrane
De nombreux pathogènes bactériens et viraux se sont dotés de mécanismes
pour échapper à l’action du complément
3-7 La reconnaissance des microbes par les phagocytes 68
La phagocytose est un mécanisme majeur de destruction des microbes
Les phagocytes ont des récepteurs pour la reconnaissance et l’internalisation
des microbes
3-8 Les mécanismes de la phagocytose 70
La phagocytose s’effectue suivant trois processus distincts
La phagocytose des cellules apoptotiques est généralement anti-inflammatoire
Des récepteurs inhibiteurs peuvent réguler la phagocytose
3-9 Les mécanismes de destruction mis en jeu par les phagocytes 72
Les phagocytes disposent de nombreux mécanismes pour tuer les microbes
internalisés
Des formes très réactives d’oxygène et d’azote sont utilisées pour tuer
les micro-organismes internalisés
3-10 La famille de récepteurs de type Toll de l’immunité innée 74
La reconnaissance de composants conservés des microbes par les récepteurs
de type Toll mène à l’inflammation et à l’activation des cellules immunitaires
sentinelles
Des molécules accessoires facilitent la reconnaissance de certains ligands
par les récepteurs de type Toll
Les récepteurs de type Toll des mammifères reconnaissent leurs ligands à la surface
cellulaire ou à l’intérieur de la cellule
3-11 Les récepteurs de type Toll donnent un signal de production de cytokines 76
Les récepteurs de type Toll des mammifères activent la production
de cytokines pro-inflammatoires et d’interféron
Les récepteurs de type Toll des mammifères utilisent deux voies
de signalisation importantes
3-12 Détecteurs intracellulaires de composants du peptidoglycane bactérien 78
NOD1, NOD2 et la cryopyrine sont des détecteurs intracellulaires
de sous-structures du peptidoglycane
Le domaine NOD génère le signal induit par les ligands
3-13 L’inflammation : déclenchement d’une réponse inflammatoire 80
La reconnaissance immunitaire innée déclenche une réponse inflammatoire
qui concentre l’action du système immunitaire sur les foyers infectieux
Les cytokines et les médiateurs lipidiques sont d’importants inducteurs
de l’inflammation
3-14 L’inflammation : recrutement de cellules immunitaires 82
L’extravasation des leucocytes s’effectue en plusieurs étapes
Les chimiokines et de petites molécules chimiotactiques dirigent
les phagocytes vers les foyers infectieux
3-15 L’inflammation : régulation et effets systémiques 84
Les réponses inflammatoires comportent deux phases : la phase aiguë et la phase
dite chronique
Les virus ont développé des systèmes pour moduler les réponses inflammatoires
Les cytokines pro-inflammatoires ont une action systémique de renforcement
de l’immunité
L’inflammation est maintenue sous contrôle par des mécanismes de rétro-inhibition
3-16 La défense antivirale innée : les interférons 86
Les interférons jouent un rôle crucial dans la défense contre
les infections virales
Les interférons sont produits par les cellules de l’immunité innée
et par les cellules infectées
xvii
DeFranco_liminaires.qxp 1/28/09 1:43 PM Page xvii

3-17 Inhibition de la réplication virale par l’interféron 88
L’interféron arme les cellules pour bloquer la réplication des virus
Les virus peuvent échapper aux effets de l’interféron
Les interférons sont utilisés dans le traitement de certaines maladies
3-18 Défense antivirale innée : induction de l’apoptose 90
La réplication virale déclenche des mécanismes apoptotiques dans la cellule infectée
Les récepteurs à domaine de mort peuvent induire la mort des cellules infectées
par un virus
Les virus ont développé de nombreux mécanismes pour bloquer l’apoptose
3-19 Les défenses intracellulaires dirigées contre les acides nucléiques viraux 92
Les défenses immunitaires innées contre les virus comprennent
des mécanismes dirigés contre l’acide nucléique viral
APOBEC3G est une cytidine désaminase qui mute les génomes rétroviraux
L’interférence d’ARN est un mécanisme de défense antivirale chez les plantes
et probablements chez les animaux
CHAPITRE 4 L’immunité acquise et la détection
de l’infection par les lymphocytes T
4-0 Le rôle des cellules T dans la réponse immunitaire acquise : un aperçu 96
L’immunité acquise diffère de l’immunité innée sous trois aspects
Les lymphocytes T tuent les cellules infectées par des virus et activent les cellules B
et les phagocytes
Les principales populations de lymphocytes T sont définies
par les classes de molécules CMH qu’elles reconnaissent
Les cellules dendritiques assurent le couplage entre l’immunité innée
et l’immunité acquise
4-1 Les cellules dendritiques et l’induction de l’immunité acquise 98
La fonction des cellules dendritiques est de présenter l’antigène
aux cellules T naïves
Les cellules dendritiques immatures sondent activement leur
environnement tissulaire
La reconnaissance d’une infection habilite les cellules dendritiques
à activer la différenciation des cellules T naïves en cellules effectrices
4-2 La structure et la fonction des molécules CMH 100
Les molécules du complexe majeur d’histocompatibilité surveillent
les compartiments internes des cellules
Les deux classes de molécules CMH ont une architecture commune
mais des fonctions distinctes dans l’immunité
4-3 Le CMH et le polymorphisme des molécules CMH 102
Les gènes du complexe majeur d’histocompatibilité codent un grand nombre
de molécules qui exercent différentes fonctions dans l’immunité
Les molécules CMH classiques sont hautement polymorphes
4-4 Molécules CMH apparentées à la classe I 104
Les molécules non classiques du CMH de classe I sont un groupe
hétérogène de variants structuraux des molécules CMH de classe I
Les cavités de liaison des peptides des molécules non classiques
du CMH de classe I sont adaptées à différentes fonctions spécialisées
4-5 La liaison des peptides par les molécules CMH 106
Les peptides se lient de façon différente aux molécules CMH de classe I
et de classe II
Des poches variables dans la cavité de liaison des peptides servent d’ancrage
à des chaînes latérales spécifiques des peptides
4-6 La présentation de l’antigène par les molécules CMH de classe I 108
Les molécules CMH de classe I présentent des peptides générés dans le cytosol
Des complexes enzymatiques dans le cytosol dégradent les protéines en peptides
Il existe dans le réticulum endoplasmique une machinerie codée principalement
dans le CMH et qui charge les peptides sur les molécules CMH de classe I
4-7 La présentation de l’antigène par les molécules CMH de classe II 110
Les molécules CMH de classe II présentent des peptides dérivés de pathogènes
internalisés par des cellules immunitaires spécialisées
Les molécules CMH de classe II récupèrent des peptides antigéniques
dans les voies de dégradation endosomiques
xviii
DeFranco_liminaires.qxp 1/28/09 1:43 PM Page xviii
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%