Anatomie interne de véligères planctoniques de

1
Découvrez plus de documents
1
accessibles gratuitement dans Archimer
Art.
NO
260
Contribution
COB
No
191
THALASSIA JUGOSLAVICA
10
i(1142)
3719-399
(1974)
Conf erence Paper
Anatomie interne
de
véligères planctoniques de Prosobranches
Mésogastropodes au stade proche de la métamorphose*
C.
Thiriot-Quiévreux
Centre Océanologique de Bretagne
-
B.
P,
337
-
29273
Brest
-
France**
-
Avec les techniques histologiques utilisées gour l'étude des
larves d'Hétéropodes13,
14,
les structures histologiques des larves
planctoniques sont décrites au stade proche de la métamorphose;
les espèces sont choisies parmi les familles de Mésogastropodes
(Lamellariidae, Eratoidae, Cypraeidae, Naticidae) dont la lignée phy-
logénique est proche de la superfamille des Atlantacea
(=
Hetero-
poda) d'après la classification de Taylor et Sohli5. Le manteau, la
cavité palléale, le pied et le tube digestif sont plus particulièrement
décrits.
-
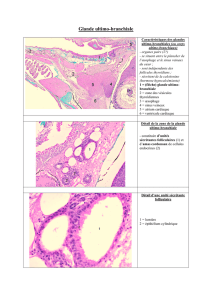
C'est au niveau du manteau et de la cavité palléale que les
différences interspécifiques sont les plus nettes. Le bourrelet pal-
léal, constitué de deux assises épithéliales, est plus ou moins dé-
veloppé
et
mo,ntre des variations !danils la nabure et la quantité des
sécrétions. Chez Velutina velutina, la glande hypobranchiale est
formée d'un bpithélium glandulaire très haut, où certaines cellules
montrent une accumulation du produit de sécrétion I(mucopo~y-
saccharide acide) qui a la forme d'un ellipsoïde et se dévesse dam
la cavité palléale. Les extrémités du manteau sont libres et dé-
bordent dle pairt et d'autre de l'ouverture de la coquille chez Erato
voluta et Lamellaria ap.; chez Cypraea sp., le manteau déborde sous
le pied seulement. Les filaments branchiaux sont caractériisés chez
Natica alderi par la présence de mucocytes
à
leurs extrémi'tés.
Le schéma général du tube digestif est remarqua~blment con-
stant d'une espèce
à
l'autre et conzprend: l'oesophage avec me
partie orale
à
mucocytes, un complexe buccal avec les ébauches
du
proboscis, de la gaine radulaire, des cartilages radulaires et des
glandes salivaires, présentes ou non selon les espèces; l'estomac
avec un bouclier ga~strique et lui faisant suite un »caecum« du sty-
let
(=
»style sac«) avec dans sa lumière un protostyle; une courte
zone de cellules allongées
à
grains APS-positifs marquant le début
de l'intestin; l'intestin; la glande digestive composée de
2
lobes avec
2
catkgories cellulaires (cellules principales et cellules
à
ergasto-
plasme) chez touttes les espèces ex'aminées sauf chez Cypraea sp.
où une troisième catégorie cellulaire est caractérisée par la présen-
ce de concrétions de teinte ocre naturelle.
Au cours de la discussion, la comparaison des résulta~ts obtenus
permet d'ébaucher l'organisation histologique type d'une véligère
planctonique de Mesogastropoda tout en mettant en évidence des
variations structurales spécifiques.
*
Contribution no
191
du Département Scientifique du Centre Océanologique de
Bretagne.
**
Adresse actuelle: Station Zoologique,
06230
-
Villefranche-sur-Mer, France.
-
17
-
Découvrez plus de documents
a
ccessibles gratuitement dans Archimer

C.
THIRIOT
-
QUIÉVREUX
1.
INTRODUCTION
Le développement et la morphologie externe des larves de Mollusques
Prosobranches ont fait l'objet de nombreux travaux, par contre leur anatomie
interne est peu connue, particulièrement en ce qui concerne l'histologie de
l'organogenèse larvaire. Je rappellerai ici brièvement les travaux réalisés sur
ce sujet depuis le mémoire de Franc1 dont la bibliographie regroupe les travaux
antérieurs.
Franc1 étudie le développement de Pisania maculosa, espèce
à
développe-
ment direct,
à
l'aide d'observations macroscopiques et microscopiques, l'embry-
ologie est décrite ainsi que l'histologie des principaux organes larvaires au
cours du développement; des observations sur le développement d'autres espèces
méditerranéennes sont données, Pisania d'orbignyi, Columbella rustica, C. scrip-
ta, Ocinebra aciculata, Purpura haemostoma, Mitra ebenus et Conus rnediter-
raneus.
Wernervécrit l'anatomie de la véligère et
de
la véliconche de Crepidula
fornicata
à
l'aide d'observations
à
travers la coquille. Lors d'une étude exhaus-
tive de l'embryologie des Mollusques, Fioroni3 observe la structure histologique
des larves
de
nombreuses espèces dont la majorité ont un développement intra-
capsulaire, quelques esp&ces mentionnées ont un développement
à
phase péla-
gique (Nassa reticulata), mais seule la véligère
à
l'éclosion est étudiée; 1' auteur
présente une bibliographie importante sur l'embryologie des Mollusques. Fio-
roni et Portmann4 décrivent la morphogenèse du tube digestif de Fusus, qui
éclôt au stade benthique.
D'Asaro5 met au point des techniques d'élevage chez des es?èces
à
déve-
loppement
à
phase pélagique et étudie chez Strombus gigas le dévelopsement des
principaux organes au cours de la vie larvaire jusqu'au 606me jour après
l'éclosion; l'étude histologique est menée sur coupes
à
la paraffine tous les 10
p
et après une coloration
à
l'éosine-hématoxyline. Avec la même technique,
D'Asaro6* étudie l'embryologie et l'organogenèse au stade proche de I'éclosion
(10ème jour après) chez Thais haemostoma, Bursa corrugata et Distorsio clat-
hrata.
Atkinsons décrit brièvement la structure histologique d'une larve de Nassa-
rius obseletus âgée de
9
jours.
Avec
des
observations
à
travers la coquille, Fretter et Grahamg présentent
des schémas de l'organisation larvaire de Lamellaria perspicua et Trivia mo-
nacha, mais sans préciser leurs structures histologiques. Fretterl0*l1 et Fretter
et Montgomery12 apportent des renseignements sur la ciliature et le mouvement
des aliments
à
l'intérieur de la larve.
La plupart de ces travaux mentionnent des espèces
à
développement intra-
capsulaire, seul D'Asaro5 apporte des renseignements importants sur des espèces
à
développement
à
phase pélagique, mais les techniques histologiques mises en
oeuvre sont peu variées.
L'organogenèse d'une espèce holoplanctonique, Atlanta lesueurils est décrite
jusqu'à la métamorphose comprise
à
l'aide de techniques histologiques compre-
nant des colorations topographiques variées et quelques réactions histochimi-
ques; les larves de trois autres genres d'Hétéropodes: Carinaria, Pterotrachea
et Firoloida sont étudiées de
la
même manière14 au stade proche de
la
méta-
morphose.

VÉLIGÉRES DE PROSOBRANCHES MÉSOGASTROPODES
Dans ce travail, utilisant les mêmes techniques que pour l'étude larvaire
des Hétéropodes, je décrirai les structures histologiques de larves planctoniques
au stade proche de la métamorphose et pendant la métamorphose, lorsque
celle-ci aura été observée. Les espèces sont choisies 'parmi les familles de Meso-
gastropoda dont la lignée phylogénique est proche de celle des Atlantacea
(=
Heteropoda) d'après la classification de Taylor et Soh1l5:
Velutina velutina Müller, Lamellariacea, Lamellariidae;
Lamellaria sp.
(L.
perspicua
4-
L.
latens, Lamellariacea, Lamellariidae);
Erato voluta (Montagu), Lamellariacea, Eratoidae;
Cyp~aea sp., Cypraeacea, Cypraeidae;
Natica alderi
(=
N.
nitida) Forbes, Naticacea, Naticidae.
Les organes du complexe palléal, le pied et le tube digestif sont plus par-
ticulièrement étudiés, les autres organes seront indiqués seulement sur les
illustrations. La comparaison des résultats permettra d'ébaucher l'organisation
histologique type d'une véligère planctonique de Mesogastropoda t,out en met-
tant en évidence des variations structurales spécifiques.
II. TECHNIQUES
Les véligères ont été récoltées dans le plancton de la région de Banpls-sur-
-Mer* (Velutina, Lamellaria, Erato et Natica) et au cours d'une campagne océano-
graphique de 1'Atlantis II (A II, 59, Woods-Hole Qceanographic Institution) en Mé-
diterranée Orientale (Cypraea).
Après avoir été triées et isolées du plancton, les larves sont fixées par les liquides
de Halimi, Bouin, Stieve ou Baker.
L'étude topographique
a
&té réalisée sur coupes sériées de
4 à
5
p
colorées par
l'azan de Heidenhain, le trichrome de Masson ou la fuchsine paraldéhyde après oxy-
dation par le mélange de Gomori.
Quelques réactions histochimiques sont été effectuées:
-
Caractérisation des composés APS-posi'tifs et notamment des glucides:
réaction
à
l'acide périodique-Schiff suivie de coloration
à
l'hématoxyline de Groat
ou au bleu de toluidine
à
pH
4,5.
-
Caractérisation des mucopolysaccharides acides: réaction métachromatique
au bleu de toluidine ou coloration au bleu alcian
à
pH
3,2
selon Mowry suivi de
réaction
à
1'APS et d'une coloration
à
l'hématoxyline.
-
Caractérisaltion des protéines: réaction
à
l'alloxane-Schiff chez Cypraea, Na-
tica alderi et Velutina velutina.
-Caractérisation des lipides héterophasiques: coloration alu noir Soudan de
matériel fixé au Balker et postchromé (chez Natica alderi) et noircissement osmique
sur coupes semi-fines après fixation selon André modifiéte, chez Velutina et Natica.
Les résultats positifs aprk application de ces deux méthoides rendent proba~bles la
présence de lipides.
-
Caractérisation des mélanines par blanchiment
à
l'eau oxygénée chez Natica
alderi.
III. DESCRIPTIONS HISTOLOGIQUES
1.
Velu'tina velutina,
Lamellariidae (larve échinospire nautiloïde)
La larve échinospire de cette espèce présente une coquille avec
5
larges
denticulations près de son ouverture et une échinospire gl~~buleuse, qui, sur
coupes histologiques, est légèrement teintée par le bleu alcian et contient dom
des mucopolysaccharides acides.
*
Ces récoltes ont
été
effectuées au Laboratoire Arago de
196b
à
1972.

C.
THIRIOT
-
QUIBVREUX
PLANCHE
1
Larves
de
Velutina velutina
1.
Coupe oblique de la partie antérieure (APS-bleu toluidine,
x
145); remarquer le pied et
l'ouverture de la glande moqueuse pédieuse antérieure (flèche).
2.
Coupe sagittale (APS-bleu toluidine,
x
115).
3.
Zone antérieure de l'oesophage (bleu alcian-APS-hématoxyline de Groat,
x
285).
4.
Coupe longitudinale de la zone pédieuse (APS-bleu toluidine,
x
285); remarquer l'&pi-
thélium glandulaire, les glandes muqueuses antérieure et postérieure (fléches).
5.
Détail de l'ébauche de la glande salivaire (flhche) (bleu alcian-APS-hématoxyline de
Groat,
x
285).
6.
Detail de la glande hypobranchiale (APS-bleu toluidine,
x
1100), noter l'écoulement du
produit de sécrétion dans la cavité palléale.
BP,
bourrelet palléal; C, coquille larvaire; Cr, cartilage radulaire; Cs, »caecum« du stylet;
E,
estomac; Ec, échinospire; GD, glande digestive; Ggo, gan$lioi~ osphradial; Ggp, ganglions
pédieux; Gr, gaine radulaire;
Hyp,
glande hypobranchiale;
1,
intestin;
Oe,
oesophage;
P,
pied;
R,
radula; S, statocyste;
T,
tentacule;;
V,
vélum.

Le bourrelet palléal:
Le bourrelet palléal est constitué de deux assises épithéliales, l'assise in-
férieure a des cellules plates et de petite taille avec quelques rares grains APS-
-positifs; l'assise supérieure
a
des cellules hautes, souvent de forme ovoïde, leur
noyau est basal avec une chromatine dis2osée en grains fins et un gros nucléole,
leur cytoplasme est rempli dans sa partie apicale de sécrétion qui se colore selon
les cellules soit
à
1'APS soit au bleu alcian, mettant en évidence la présence de
polysaccharides, dont la nature reste
à
préciser, et de mucopolysaccharides
acides; dans certains cas, le cytoplasme montre des travées qui lui donnent un
aspect alvéolaire caractéristique.
L'osphradie:
Le ganglion osphradial et l'osphradie forment un renflement dans la cavité
palléale et ne présentent )pas de formation glandulaire. Les filaments branchiaux
sont nettement individualisés sur la cténidie.
La glande
h
ypobranchial e:
Dans le plafond de la cavité palléale, en forme de
U,
entre la cténidie et le
rectum s'étend la glande hy>obranchiale, ou épithélium glandulaire, très déve-
loppée chez cette espèce. Les cellules, larges et hautes, présentent des noyaux
avec un gros nucléole et une chromatine disposée en grains, une zone riche en
ergastoplasme est parfois visible
à
la base de la cellule après une coloration au
bleu de toluidine; le cytoplasme est rempli de dcrétion se présentant sous
plusieurs aspects selon l'activité de la cellule: soit en grains très fins, cyano-
philes, colorés par la fuchsine paraldéhyde après oxydation, faiblement positif
au bleu alcian; soit en flaque ayant parfois la forme d'une ellipsoïde se colorant
intensément de la même manière que précédemment, cette sécrétion présente
les caractéristiques des rnucopolysaccharides acides. D'autres cellules montrent
des grains ovales plus ou moins coalescents positifs
à
I'APS et
à
l'alloxane43chiff
et donc de nature protéique. Parfois la cellule est vide de tout produit de
sécrétion et le cytoplasme paraît hyalin. Entre ces cellules glandulaires s'obser-
vent des cellules étroites et de petite taille qui ont un noyau disposé soit latéra-
lement soit dans la partie apicale.
L'épithélium tégumentaire de la région céphalique est simple et sans sécré-
tion figurée.
Le pied:
Le pied est revêtu d'un épithélium qui, au niveau de la sole, se compose
de hautes cellules ciliées glandulaires, les noyaux sont basaux avec une chro-
matine dense, les cytoplasmes sont remplis de sécrétion qui, selon les cellules,
se présente soit sous la forme de grains de nature glucidique, soit en flaque
diffuse ayant les propriétés des mucopolysaccharides acides. Une zone de cellu-
les
à
cytoplasme fortement APS-positif s'étend sous l'épithélium glandulaire et
correspond
à
la glande muqueuse pédieuse antérieure. La glande muqueuse
pédieuse postérieure est située vers la partie basale du pied, près de l'extrémité
des ganglions pédieux, et s'ouvre au milieu de la base du pied. La plupart des
cellules sont larges et hautes avec une sécrétion constituée de mucopolysaccha-
rides acides, mais quelques cellules étroites s'intercalent entre les mucocytes
et montrent une sécrétion APS-positive.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%