introduction - SVT au Lycée Maupassant

introduction
En première S, nous avons montré que des mutations (spontanées ou liées à l’environnement) pouvaient être à l’origine de nouveaux
allèles et donc en partie expliquer une diversité des êtres vivants. Dans le chapitre 1A1, nous avons vu comment les mécanismes
impliqués dans la reproduction sexuée, participent au brassage des allèles et favorisent la diversité au sein des espèces.
Ainsi avec les connaissances acquises à ce jour :
Dans ce chapitre 1A2, nous allons voir que d’autres mécanismes (impliquant ou non des modifications du génome) contribuent à la
diversification des êtres vivants.
Le chapitre 1A3 permettra de montrer comment les mécanismes de diversification (liés à la reproduction sexuée (chap. 1A1) ou non
(chap.1A2)), participent à l’évolution de la biodiversité notamment à l’échelle de la diversité des espèces (spéciation).
?
Chap. 2
BRASSAGE des allèles lors de la reproduction
sexuée
(Méiose x fécondation) : Chap.1A1
Mutations somatiques ou germinales à
l’origine des différents allèles d’un gène :
DIVERSITE GENETIQUE DE L’ESPECE
DIVERSITE des individus d’une espèce.
Est à l’origine de …
?
Chap.1

Atelier 1 : Hybridation et polyploïdisation chez les Salamandres
Une grande diversité parmi les Salamandres d’Amérique du Nord
Le document 1 décrit 4 espèces de salamandre reconnaissables à leurs caractéristiques morphologiques (taille, couleur…).
Le document 2 indique que parmi les Salamandres de Jefferson, on a repéré différentes populations :
- Certaines populations sont constituées d’individus mâles et femelles dont le caryotype est diploïde. (2n chromosomes) et
qui pratiquent une reproduction sexuée « classique ».
- D’autres populations ne sont constituées que d’individus femelles difficiles à différencier des populations diploïdes (à part
la taille un peu plus importante des individus) et dont le caryotype peut présenter une ploïdie variable en fonction de la
population : 3n ; 4n ; 5n. De plus les chromosomes constituant ces caryotypes proviennent d’un mélange variable d’un
individu à l’autre entre les 4 espèces pures présentées dans le document 1.
Il existe une diversité des caryotypes (en nombre et en variétés des lots de chromosomes) au sein des Salamandres de Jefferson.
Origine de la diversification des Salamandres
Le mélange de lots de chromosomes appartenant à des espèces différentes de Salamandre présentées dans le document 2 permet
de penser à un mélange = une hybridation entre des individus d’espèces différentes.
Les œufs de ces femelles hybrides ne subissent pas la méiose et possèdent donc un nombre de chromosomes identique aux cellules
somatiques de la mère.
Si la mère est diploïde, l’œuf est diploïde. Le spermatozoïde du mâle (issu des populations de salamandres diploïdes « pures » grâce
à la proximité géographique (cf doc 1), s’il est indispensable au développement de l’œuf
- peut ne pas être incorporé à l’œuf, la salamandre reste alors diploïde.
- peut être incorporé en échange d’un lot maternel, la salamandre reste aussi diploïde.
- peut être incorporé à l’œuf, la salamandre est alors triploïde.
Le même processus permet de passer à des états de ploïdie supérieur si la mère est triploïde ou tétraploïde.
C’est l’hybridation suivi de la polyploïdisation au cours de la kleptogenèse qui permet de diversifier les génomes des
salamandres.

Atelier 2 : Gènes du développement
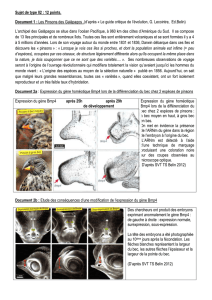
Doc 1 : Malgré des formes très variées le plan d’organisation des êtres vivants est sous le contrôle d’une famille de gènes
homologues (les gènes Hom pour les invertébrés et Hox pour les vertébrés)
Doc 2 : Le gène HoxD13 s’exprime d’une manière plus durable et dans une zone plus étendues lors du développement de l’embryon
de souris que lors du développement de l’embryon d’un poisson zèbre. ce qui aboutit à la formation d’un membre supérieur
typique des vertébrés tétrapodes (humérus, radius-cubitus, doigts) alors que chez le poisson l’expression est plus limitée en surface
et dans le temps aboutissant à la mise en place d’une nageoire.
La mise en place du membre supérieur d’un poisson et d’une souris n’est pas liée à des gènes différents. Les différences ont pour
origine le lieu et le temps d’expression du gène HoxD13.
Doc 3 : Les pinsons des iles Galápagos ont des becs très divers liés à leur nutrition (petit bec pointu insectivore, gros bec
granivore…)
La formation du bec se fait à l’état embryonnaire, et est sous le contrôle d’un gène nommé Bmp4. Ce gène présente d’importante
similitude chez tous les types de pinsons. C’est l’expression de ce gène au niveau de la tête qui aboutira aux différents becs.
Si le Bmp4 est sous exprimé le bec sera petit , surexprimé il sera épais et crochu et exprimé « normalement » il sera moyen et
pointu.
Conclusion : Ainsi les variétés de phénotypes ne dépendent pas que des variétés alléliques mais reposent aussi sur la foçon dont un
même gène s’exprime lors du développement. De ce faite les gènes du développement par leur fonctionnement participe à la
diversité des forme de vie.
Complément :
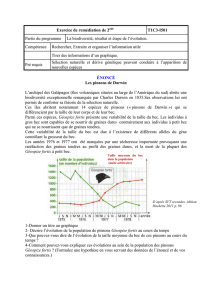
Lors de son voyage au Galápagos, Darwin a découvert une diversité importante de pinsons. Ces derniers différaient notamment par leurs tailles et
la forme de leurs becs. Darwin établit un lien entre la forme des becs des oiseaux et le régime alimentaire de ces derniers. Il expliqua la diversité
des pinsons sur les différentes îles, par une diversification à partir d’une même espèce ancestrale. Ainsi en fonction des ressources alimentaires
présentes sur les îles les pinsons ayant le bec le plus adapté ont survécu. Au cours des générations et aléatoirement, certaines modifications du
bec se produisent. La ressource alimentaire restreinte (pression de sélection du milieu) sélectionne les oiseaux dont le bec et le plus adapté
(variable selon les îles). C’est le mécanisme de sélection naturelle.
Darwin n’avait aucune idée de la notion de gène. Il ne pouvait donc pas savoir quels « éléments » du pinson étaient modifiés aléatoirement. On sait
aujourd’hui que ce sont les gènes (mutations génétiques) qui peuvent être modifiés au hasard.
Sélection naturelle= Sélection par l’environnement d’un l’allèle apportant un avantage (ou évitant un désavantage) à l’individu qui le porte. Les
individus favorisés se reproduisent plus et transmettent donc plus l’allèle sélectionné aux générations suivantes.
Dans le cas du bec des pinsons, c’est le gène Bmp 4 qui peut être modifié par mutation. Ce gène intervient lors du développement embryonnaire
(gène de développement), et peut être soit surexprimé soit sous exprimé.
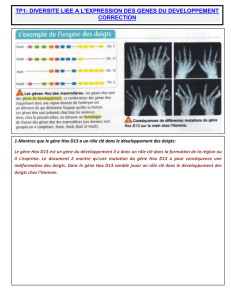
Pour démontrer le lien entre Bmp4 et la forme du bec on peut étudier les expériences de transfert de gène réalisée en 2004. En transférant dans
les cellules embryonnaires frontales de poulet un gène Bmp4 surexprimé, on constate qu’au 10ème jour après la fécondation, l’embryon possède un
bec plus haut et plus large que le témoin. Au contraire l’introduction d’un gène sous-exprimé de Bmp4 induit une diminution de la taille du bec
par rapport au témoin (embryon au gène Bmp4 non modifié). Bmp4 est donc bien impliqué dans la mise en place du bec et sa morphologie. Selon
l’intensité de son expression le même gène aboutira à des becs différents
Les études de 2004 montrent que selon les espèces de pinsons, l’expression du gène au stade 4,5 jours du développement est variable. Les espèces
ayant à l’âge adulte un bec fin (Geospiza difficilis, Geospiza scandens) n’expriment pas ce gène (aucune transcription à partir du gène Bmp4). Au
contraire les espèces à gros bec (Geospiza fortis ou Geospiza magnirostris) expriment fortement le gène.
La diversité de bec dépend donc du timing de l’expression du gène Bmp4.
Au stade 5,5 jours toutes les espèces expriment Bmp4, mais à des degrés diverses. Les espèces à gros bec l’expriment très fortement et dans de
nombreuses cellules embryonnaires du futur bec. Au contraire les espèces à petit bec expriment faiblement Bmp4 et dans peu de cellules.
Selon le degré d’expression et la localisation de l’expression de Bmp4, le bec formé à une morphologie variable.
Conclusion :
Darwin avait mis en évidence la diversification des espèces de pinson à partir d’une espèce ancestrale par sélection naturelle (les pinsons ayant un
bec plus adapté aux ressources alimentaires de l’île étant favorisés).
On sait aujourd’hui que les variations de la forme du bec sont liées au timing et degré d’expression du gène Bmp4 (gène architecte). Ainsi une
modification aléatoire de Bmp4 dans une population de pinson, peut conduire à une nouvelle forme de bec, qui sera ou non avantageuse. On
explique génétiquement la sélection naturelle de Darwin.

Atelier 3 : Polyploïdies
Doc 1 : Des croisements sont possibles entre des espèces différentes de fraisiers. Ces croisements sont appelés hybridation. Dans le
cas du fraisier l’hybride obtenu entre chiloensis et virginiana possède un caryotype octoploide c'est-à-dire 8 chromosomes par
« paire » au lieu de 2 habituellement.
Un hybride cumule les génotypes de ces parents et présente donc les caractéristiques des deux parents ce qui permet en
agriculture d’additionner les caractéres positif pour l’homme de plantes (ici le bon gout et la taille)
Doc 2 : un individu polyploide possède plus de 2 chromosomes homologues. Ce phénomène courant chez les végetaux est rare chez
les animaux , il est lié à des anomalies lors de la mitose soit thermique ou chimique ou à des anomalies lors de la méiose.
Doc 3 : Le caryotype de F virginiana montre 4 chromosomes homologues par « paire » ces cellules sont donc tétraploide 4n=28
Doc 4 : Les deux espéces originelles osnt très différentes par la taille et le gout l’hybride possede la taille et le gout des 2 « parents »
Doc 5 : La colchicine bloque l’anaphase de mitose, les chromatides se séparent mais restent dans la même cellule. A la fin de cette
mitose la cellule possède donc 4 jeux de chromosomes homologues identiques 2 à 2. Si cette division anormale a lieu au début du
développement du fraisier elle pourra être conservée dans l’individu adulte si les autres mitoses se déroulent normalement.
Conclusion : Des anomalies lors de la mitose ou de la méiose peuvent aboutir à des cellules polyploides possédant plus de 2
chromosomes homologues.
Voir aussi brassicacées (allo et autoploïdie)
Correction Atelier 3 Brassicacé
Doc 2 : Chez les plantes, il existe des cas de croisements entre espèces voisines (allopolyploïdisation) qui dans de rares
cas peuvent produire des plantes fertiles et conduire à de nouvelles espèces. Ainsi chez les Brassicacées, les trois
espèces B. napus, B. juncea et B. carinata, dérivent, respectivement, du croisement entre B. rapa et B. oleracea, B. rapa
et B. nigra, B. nigra et B. oleracea comme illustré dans la figure ci-dessous.
Croisements interspécifiques chez les Brassicacées
Doc 3 : L’allopolyploïdisation est un cas de polyploïdisation qui correspond au rassemblement de génomes diploïdes
différents (hybridation inter-spécifique). Les hybrides interspécifiques ont une fertilité très limitée puisque les
chromosomes ne sont présents qu’en un seul exemplaire et ne peuvent donc s’apparier pour la méiose. La duplication
du génome lors d’une méiose anormale restaure la possibilité d’appariement des chromosomes homologues et donc la
fertilité.

Conclusion : Chez les plantes , la spéciation peut se produire de façon assez rapide suite à la duplication du génome
(polyploïdie) qui intervient généralement par formation de gamètes non réduits. Les autopolyploïdes se forment par
duplication du génome d’une même espèce tandis que les allopolyploïdes résultent de la duplication du génome d’un
hybride interspécifique.
Doc 4 : L’autopolyploidisation est un cas de
polyploidisation qui correspond au rassemblement
de génomes diploïdes semblables (hybridation intra-
spécifique). Les autoploïdes se forment par
duplication du génome de la même espèce, le plus
souvent lors d’une non-réduction gamétique
(erreur de déroulement de la méiose).
 6
7
8
6
7
8
1
/
8
100%