qwertyuiopasdfghjklzxcvbnmqwerty - ORBi

qwertyuiopasdfghjklzxcvbnmqwerty
uiopasdfghjklzxcvbnmqwertyuiopasd
fghjklzxcvbnmqwertyuiopasdfghjklzx
cvbnmqwertyuiopasdfghjklzxcvbnmq
wertyuiopasdfghjklzxcvbnmqwertyui
opasdfghjklzxcvbnmqwertyuiopasdfg
hjklzxcvbnmqwertyuiopasdfghjklzxc
vbnmqwertyuiopasdfghjklzxcvbnmq
wertyuiopasdfghjklzxcvbnmqwertyui
opasdfghjklzxcvbnmqwertyuiopasdfg
hjklzxcvbnmqwertyuiopasdfghjklzxc
vbnmqwertyuiopasdfghjklzxcvbnmq
wertyuiopasdfghjklzxcvbnmqwertyui
opasdfghjklzxcvbnmqwertyuiopasdfg
hjklzxcvbnmrtyuiopasdfghjklzxcvbn
mqwertyuiopasdfghjklzxcvbnmqwert
yuiopasdfghjklzxcvbnmqwertyuiopas
dfghjklzxcvbnmqwertyuiopasdfghjklz
xcvbnmqwertyuiopasdfghjklzxcvbnm
qwertyuiopasdfghjklzxcvbnmqwerty
Aide aux Cliniques de pédiatrie :
éléments de:
bactériologie et virologie
Ulg, faculté de médecine
Professeur Oreste Battisti

Introduction
Le diagnostic des affections pédiatriques repose sur les éléments classiques :
l’anamnèse, l’examen clinique et les données paracliniques.
En ce qui concerne les données paracliniques, l’imagerie et la microbiologie seront souvent
utilisées ( voir « Maladies infectieuses de l’enfant et immunologie » et « Chirurgie et imagerie
de l’enfant »).
Nous faisons dans cet ouvrage un rappel utile des méthodes microbiologiques ainsi qu’une
présentation intégrée à la clinique, des microorganismes potentiellement responsables des
affections dues à une infection bactérienne ou virale.
Nous remercions les collègues microbiologistes pour leur contribution.


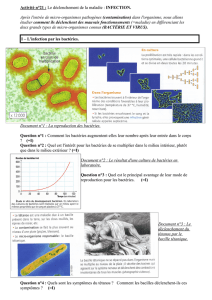
A PROPOS DU DIAGNOSTIC D'UNE INFECTION BACTERIENNE
A - GÉNÉRALITÉS
Le diagnostic bactériologique est un ensemble de moyens permettant de
confirmer telle ou telle étiologie infectieuse d'origine bactérienne. Ces moyens
diagnostiques sont variés et caractérisent soit le diagnostic direct soit celui
indirect :
DIAGNOSTIC DIRECT : mise en évidence de la bactérie elle-même, donc
finalement de sa culture ou isolement qui permettra l'identification ultérieure
ainsi que de préciser sa sensibilité aux antibiotiques (antibiogramme).
DIAGNOSTIC INDIRECT : mis en évidence de la réponse de l'organisme à
l'infection par la présence d'anticorps spécifiques, le plus souvent sériques ou
plus rarement par une réponse d’hypersensibilité, dite allergique.
B - DIAGNOSTIC DIRECT
La prescription s'intitulera "examen cyto-bactériologique" d'une urine (ECBU),
d'une expectoration (ECBC), d'un liquide pleural... ayant pour objectif de
rechercher la mise en évidence de la bactérie responsable de l'infection.
1 - Demande :
Il sera important de bien identifier le patient par son nom, son prénom, sa date
de naissance, le sexe. Il existe une procédure standard de recherche de cellules
et de germes, d'où l'appellation suivante : "Examen cytobactériologique des
urines ". Celle-ci exclut toute recherche systématique de germes particuliers
telle la recherche de BAAR (Bacilles Acido-Alcoolo-Résistants), de
leptopsires...
Le clinicien ne devra jamais oublier d'indiquer toute demande ou
recherche particulière, en raison de l'utilisation de milieux spéciaux.
Exemple : La culture du bacille tuberculeux (BAAR) se fera sur le milieu de
Lowenstein-Jensen après, au minimum 3 semaines d'incubation à 37°C, le
milieu étant protégé de la dessication par un bouchon en caoutchouc.
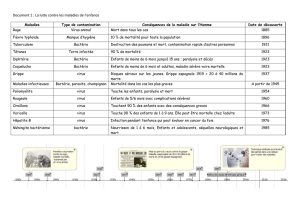
2 - Examen macroscopique :
Toute infection bactérienne s'accompagne, outre la présence de bactéries,
de signes biologiques liés à l'inflammation avec l'éventuelle présence de
leucocytes, notamment de polynucléaires.
Ces éléments peuvent entrainer au delà d'un seuil, une modification

visuelle, clairement perceptible à l'oeil nu, qui signe une anomalie
patente.
Divers éléments sont alors obtenus comme le montrent les exemples
suivants:
Trouble : urine, LCR, liquide pleural ou articulaire
Hématurique : urine, LCR,
liquide pleural ou articulaire
Autre coloration anormale
Odeur : on notera celle
caractéristique lors d'infections
à germes anaérobies stricts dans
un liquide pleural
Consistance : Exemple d'une
selle diarrhéique.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
1
/
221
100%