cellule

UCC 4 : LIVRET : LA CELLULE
gt
BPREA agri/viti
UCC G2 : BIOLOGIE
LIVRET : La cellule
GÉNÉRALITÉS
La cellule constitue l'unité structurelle et fonctionnelle fondamentale des êtres vivants. Les formes
de vie les plus simples sont représentées par des cellules uniques capables de se reproduire par
duplication. Les organismes supérieurs - c'est-à-dire presque toutes les plantes et les animaux, y
compris l'homme - sont constitués de nombreuses cellules organisées en systèmes complexes qui
ont des fonctions spécialisées et sont reliés entre eux par des systèmes de communication
sophistiqués.
PROCARYOTES ET EUCARYOTES
Le monde des êtres vivants se divise en deux grandes branches selon le type de cellules qui les
constituent. La première branche est celle des Procaryotes (du grec protos, primitif, et karion,
noyau), qui comprend les organismes unicellulaires les plus simples, c'est-à-dire les Bactéries et les
Algues bleues, dépourvus de membrane nucléaire : le matériel génétique n’est pas enfermé dans un
noyau cellulaire mais est librement immergé dans le cytoplasme.
Le cytoplasme d'une bactérie ne contient ni
organites ni noyau cellulaire. Le
chromosome de la bactérie, constitué
d'ADN, est libre dans le cytoplasme, dans
lequel « flottent » également les ribosomes.
Ce cytoplasme est entouré, comme
n'importe quelle cellule, par une membrane
plasmique, constituée de phospholipides.
Autour de cette dernière se trouve toujours
une paroi peptidique, plus ou moins épaisse.
C'est l'épaisseur de la paroi qui détermine
la réaction à la coloration de Gram, qui
permet de classer les bactéries en gram +
(paroi épaisse) et gram - (paroi fine). Une
troisième couche protège encore la cellule bactérienne : la capsule. De nombreuses bactéries
possèdent, en outre, des excroissances diverses, comme des pili (cils) et un ou plusieurs flagelles
servant à la propulsion.
D’après l’Encyclopédie Microsoft ® Encarta ® 2004.
Quand il y a abondance de substances nutritives, une seule cellule procaryote se divise toutes les
20 minutes, donnant naissance de la sorte à 5 milliards de cellules (à peu près autant que la
population humaine sur la Terre) en moins de 11 heures. Cette vitesse de reproduction permet aux
Bactéries de s'adapter rapidement aux changements du milieu.
Richard GRATTON Page 1 date de mise à jour 25/09/2011

UCC 4 : LIVRET : LA CELLULE
La deuxième branche, celle des Eucaryotes (du grec eu, vrai, et karion, noyau), comprend tous les
autres êtres unicellulaires et pluricellulaires, les végétaux et les animaux, qui ont un noyau bien
défini, avec une membrane qui sépare le matériel génétique du cytoplasme.
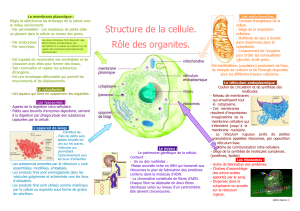
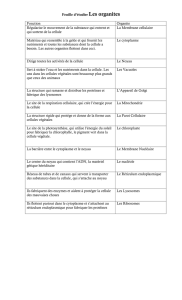
MORPHOLOGIE ET PHYSIOLOGIE DE LA CELLULE
Les cellules sont toutes semblables entre elles : leur diamètre est généralement compris entre 10 et
30 micromètres, elles sont composées des mêmes molécules de base, elles les élaborent à travers
les mêmes types de réactions chimiques et elles ont en commun de nombreux organites et des
structures identiques ou très semblables, telles que les membranes, le noyau cellulaire, les
ribosomes, l'appareil de Golgi, le réticulum endoplasmique, les mitochondries et le cytosquelette.
Par-delà cette uniformité de base, il existe des différences entre les cellules animales et les cellules
végétales. Par rapport aux cellules animales, les cellules des plantes possèdent un revêtement
supplémentaire, la paroi cellulaire, formée de cellulose. Les cellules végétales possèdent en outre
des organites caractéristiques : les vacuoles, vésicules pleines d'eau contenant un certain nombre de
substances en solution, et les chloroplastes, qui contiennent la chlorophylle et d'autres pigments.
L'eau contenue dans les vacuoles confère une certaine rigidité à chaque cellule, et assure le
maintien de la plante. C’est au sein des chloroplastes qu’est capturée l'énergie du Soleil et qu’elle
est transformée en substances énergétiques –telle que le glucose- grâce au processus de la
photosynthèse chlorophyllienne.
cellule animale cellule végétale
La membrane plasmique
Chaque cellule est enfermée dans une membrane, une enveloppe protectrice de 8-12 nanomètres
(nm), c'est-à-dire de 8-12 millionièmes de millimètre, qui délimite le compartiment cellulaire et la
sépare du milieu environnant. La membrane joue à la fois le rôle d'un filtre et d'un moyen de
transport. D'une part, elle contrôle l'entrée des substances nutritives et la sortie des déchets
cellulaires et, d'autre part, elle crée un milieu interne différent du milieu externe.
Les constituants principaux de la membrane plasmique sont les lipides (c'est-à-dire les graisses),
les protéines et, dans certains cas, les hydrates de carbone (ou sucres,
Richard GRATTON Page 2 date de mise à jour 25/09/2011

UCC 4 : LIVRET : LA CELLULE
La paroi cellulosique
Chaque cellule végétale est délimitée par une paroi située à l'extérieur de la membrane plasmique,
qui la distingue de la cellule animale plus que toute autre structure. La paroi cellulaire constitue un
abri ou une « maison » pour la cellule ; elle en détermine la forme et est responsable de sa solidité.
Le cytoplasme
L'intérieur de la cellule est rempli par le cytoplasme, une matrice aqueuse colloïdale qui contient
des organites et des systèmes complexes de membranes comme le réticulum endoplasmique et
l'appareil de Golgi. Le cytoplasme occupe à peu près la moitié du volume total de la cellule
animale (moins pour la cellule végétale à cause de la présence de la vacuole). On y trouve toutes
les substances chimiques vitales en dispersion, notamment les sels, les ions, les sucres, une grande
quantité d'enzymes et de protéines, et une grande partie de l'ARN. L'eau constitue à elle seule près
de 80 % du cytoplasme.
Le cytosquelette
Dans les cellules eucaryotes, le cytoplasme contient une sorte de châssis formé de microtubules et
de différents types de filaments et de microfilaments, qui constituent le cytosquelette. Ce dernier a
de nombreuses fonctions. C'est lui qui confère à la cellule sa forme caractéristique et la dote de
motilité en lui donnant la possibilité d'accomplir des mouvements amiboïdes. Le cytosquelette
permet en outre les déplacements des organites cellulaires et coordonne des fonctions biologiques
fondamentales, comme la division cellulaire. Dans les cellules végétales, la direction du trafic
moléculaire assurée par le cytosquelette est très importante pour la croissance de la paroi cellulaire,
car l'orientation des microfibrilles de cellulose qui sont associées à la paroi est déterminée par
l'orientation des microtubules du cytosquelette.
Les molécules et les protéines de petites dimensions sont diffusées dans le cytosol avec facilité,
comme si elles étaient plongées dans l'eau, tandis que les vésicules de transport et les organites se
déplacent très lentement et ont besoin de véritables moteurs protéiques qui, en utilisant l'ATP, les
transportent d'un côté à l'autre de la cellule. Les déplacements de ces structures cellulaires se font
grâce au cytosquelette.
Les microtubules sont des tubes creux très fins constitués d'une protéine appelée tubuline. La
cellule possède des centres d'organisation des microtubules, qui en dirigent la formation : les
centrioles, les corpuscules basaux des cils et les centromères (voir centrioles). Dans le cas des
microtubules évoluant librement dans la cellule, il existe un centre de formation principal à côté du
noyau, constitué de deux centrioles perpendiculaires l'une à l'autre (asters), à partir desquelles les
microtubules rayonnent, leurs queues négatives étant tournées vers le centre de la formation.
Les microtubules sont des structures dynamiques qui se forment et sont détruites en permanence.
Ce caractère hautement dynamique de la structure des microtubules explique toute une série de
fonctions cellulaires importantes comme, par exemple, la motilité. Le mouvement cellulaire est dû
à un réarrangement permanent du cytosquelette cellulaire.
La vacuole
Toutes les cellules végétales présentent une poche ou vésicule pleine d'eau contenant en solution
des sels et diverses substances. La membrane qui le délimite, dite tonoplaste, est responsable du
« tonus », c'est-à-dire de la tension cellulaire. À travers cette membrane, en effet, la vacuole reçoit
ou perd de l'eau, ce qui a pour effet de modifier la turgescence de la cellule végétale.
La cellule jeune se caractérise par plusieurs vacuoles de petites dimensions qui, dans la cellule
mûre, se fondent ensemble pour former une seule grande vacuole occupant 80 % du volume
cellulaire. Le cytoplasme, le noyau et les chloroplastes, avec les autres organites cellulaires, sont
ainsi relégués dans une position marginale, derrière la paroi végétale.
Richard GRATTON Page 3 date de mise à jour 25/09/2011

UCC 4 : LIVRET : LA CELLULE
La vacuole assure la croissance de la cellule végétale en absorbant de l’eau, le principal constituant
du suc qu'elle contient, car la production de nouveau cytoplasme serait trop coûteuse pour la
cellule.
Les autres constituants du suc vacuolaire, qui varient dans les différentes plantes ainsi que dans les
différents tissus d'une même plante, sont les acides aminés, les sucres, les protéines, les substances
minérales ou de réserve, les déchets et les pigments. Cette vésicule contient par conséquent les
substances les plus diverses, qui peuvent être accumulées ou mobilisées dans la cellule selon ses
nécessités de croissance ou les conditions extracellulaires.
La vacuole a souvent une action de détoxification : elle stocke des substances qui, accumulées en
trop grande quantité, pourraient nuire au cytoplasme (par exemple, les substances toxiques
destinées à lutter contre les ravages des herbivores et qui sont diffusées hors des vacuoles lorsque
l'animal s’attaque à la plante).
La vacuole contient aussi quelques pigments, dits antocyanines, responsables des couleurs bleue,
violette, rouge pourpre ou rouge foncé des fleurs, fruits et tiges. Ce sont ces mêmes pigments qui
se forment chaque année en réponse au froid, en même temps que la dégradation de la
chlorophylle, donnant aux feuilles leur coloration automnale caractéristique. Parfois, comme dans
l'érable rouge, ils sont présents dans des quantités telles qu'ils masquent la couleur verte de la
chlorophylle des feuilles.
La vacuole remplit une autre fonction importante, semblable à celle des lysosomes de la cellule
animale. Elle est capable d'englober et de dégrader des organites cellulaires vieillis comme les
ribosomes, les mitochondries ou les plastides.
Les centrioles
Les centrioles sont de petits organites cylindriques et creux, de 0,2 micron de largeur et de
0,4 micron de longueur. Chaque centriole est formé de 9 triplets de microtubules parallèles et liés
entre eux et au centre du cylindre par des bras protéiques qui forment une structure semblable à la
roue d'un char. Les centrioles se trouvent souvent par couples, disposés perpendiculairement l'un à
l'autre.
Toutes les cellules possèdent une structure appelée centrosome, constituée d'un couple de
centrioles, qui sert à organiser les microtubules du cytosquelette durant l'interphase (la période du
cycle cellulaire entre deux divisions successives) et qui se reproduit au moment de la mitose, pour
donner naissance aux deux pôles du fuseau mitotique.
Les lysosomes
Dans le cytoplasme des cellules eucaryotes sont présents des organites de forme, de nombre et de
dimensions variables, qui peuvent être considérés comme l'« estomac » de la cellule. Ce sont les
lysosomes, vésicules contenant une très haute concentration d'enzymes digestives (ou hydrolases)
qui sont utilisées pour dégrader les macromolécules.
Les lysosomes se forment et se détachent de l'appareil de Golgi, et se déplacent vers l'extérieur de
la cellule pour aller se fondre avec la membrane plasmique.
À l'intérieur des lysosomes, on trouve environ 40 enzymes hydrolytiques différentes : protéase,
nucléase, glycosidase, lipase et d'autres encore. Elles sont toutes actives dans un milieu très acide.
Étant donné que ces enzymes sont capables de digérer tous les types de macromolécules, y compris
celles qui constituent les membranes biologiques, il est très important qu'elles restent bien séparées
du cytoplasme et des autres organites de la cellule. À cette fin, la membrane des lysosomes possède
des caractéristiques particulières : elle ne peut pas être attaquée par les hydrolases, elle empêche la
diffusion de macromolécules et sélectionne les produits de la digestion qui doivent être libérés dans
le cytoplasme pour y être réutilisés.
Les lysosomes remplissent deux fonctions fondamentales : l'hétérophagie, qui est la digestion de
matériaux provenant de l'extérieur, qu'il s'agisse de substances alimentaires ou d'organismes
Richard GRATTON Page 4 date de mise à jour 25/09/2011

UCC 4 : LIVRET : LA CELLULE
pathogènes ; et l'autophagie, qui est la digestion de certaines portions de la cellule par elle-même.
Cette deuxième fonction est essentielle aussi bien dans les processus de développement, pour
lesquels il est très important de recycler certains matériaux cellulaires, que, pour assurer une vie
saine aux cellules. Ce processus permet en effet aux cellules de se débarrasser de substances
toxiques qui, en s'accumulant, pourraient l'endommager. Les cellules ou les tissus dans lesquels ce
recyclage n'a pas lieu subissent en effet une détérioration précoce.
Les mitochondries
Toutes les cellules des Eucaryotes contiennent des générateurs d'énergie extrêmement efficaces :
les mitochondries, organites de forme cylindrique de 7 microns de longueur environ, et d'un
diamètre compris entre 0,5 et 1 micron, dans lesquelles a lieu la respiration cellulaire.
Les mitochondries ont des dimensions très voisines des Bactéries. Elles sont revêtues de deux
membranes qui jouent un rôle essentiel dans le métabolisme, elles contiennent leur propre ADN et
peuvent se reproduire de façon autonome, indépendamment de la division cellulaire. En outre, elles
sont dotées d'ARN et de ribosomes qui leur permettent de synthétiser quelques protéines.
Lors de la reproduction, les mitochondries des animaux supérieurs ne se transmettent que par voie
maternelle. En effet, bien qu'elles soient présentes aussi bien dans les ovocytes (les cellules
reproductrices femelles) que dans les spermatozoïdes (les cellules reproductrices mâles), ces
derniers ne donnent que leur ADN au moment de la fécondation.
Les deux membranes qui limitent les mitochondries créent deux compartiments séparés : la matrice
interne et un espace intermembranaire, beaucoup plus petit.
La membrane externe contient un nombre élevé de copies d'une protéine appelée porine qui forme
des canaux perméables d'une taille maximum de 10 000 daltons. D’autres enzymes, participant à la
synthèse et au métabolisme des lipides, sont aussi présentes sur cette membrane.
La membrane interne est repliée en de nombreuses crêtes qui en augmentent la surface globale. La
matrice interne contient des substances importantes pour le processus de respiration cellulaire : les
enzymes, les coenzymes et les phosphates. La membrane interne contient des protéines ayant trois
fonctions différentes : certaines sont impliquées dans les réactions d'oxydation de la chaîne
respiratoire ; d'autres sont des protéines de transport spécifiques qui régulent le passage de
substances au travers de la matrice ; le dernier groupe, enfin, est constitué d'un complexe
enzymatique appelé ATP synthétase, qui synthétise l'ATP, un composé particulier qui joue le rôle
de stockeur d'énergie.
La matrice contient un concentré de centaines d'enzymes utilisées pour l'oxydation de substances
organiques et impliquées dans une séquence cyclique de réactions, connue sous le nom de cycle de
Krebs, au cours duquel de l'énergie est libérée. La matrice contient aussi de nombreuses copies
d'ADN mitochondrial, de ribosomes mitochondriaux structurellement différents des ribosomes
cytoplasmiques, d'ARN de transport, d'ARN messagers, facteurs de la synthèse protéique, et
d'enzymes nécessaires à l'expression des gènes mitochondriaux et à leur réplication (voir aussi
ADN).
Certaines caractéristiques des mitochondries, comme la présence d'un système permettant la
synthèse protéique, la capacité de s'autoreproduire, l’existence d'une double membrane, rendent ces
organites extrêmement intéressants pour l’étude de l'évolution des cellules eucaryotes. Selon une
théorie proposée par la biologiste américaine Lynn Margulis, les mitochondries auraient été à
l'origine des Bactéries aérobies, qui se servaient d'oxygène pour la respiration cellulaire. Ces
organismes auraient été incorporés dans les cellules eucaryotes à travers un processus
d'endosymbiose. En d'autres termes, certaines Bactéries libres auraient commencé à mener une vie
en association étroite avec d'autres organismes unicellulaires incorporés à leur cytoplasme. De
cette façon, la cellule hôte aurait bénéficié de l'énergie produite par la Bactérie aérobie, tandis que
cette dernière, en échange, aurait pu disposer d'une source plus efficace de substances nutritives
(voir aussi cellules plus complexes).
Richard GRATTON Page 5 date de mise à jour 25/09/2011
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%