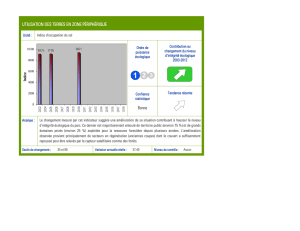
Approche quantitative de la réponse écologique des espèces

Approche quantitative de la r´eponse ´ecologique des
esp`eces v´eg´etales foresti`eres `a l’´echelle de la France
Christophe Coudun
To cite this version:
Christophe Coudun. Approche quantitative de la r´eponse ´ecologique des esp`eces v´eg´etales
foresti`eres `a l’´echelle de la France. Ecologie, Environnement. ENGREF (AgroParisTech),
2005. Fran¸cais. <tel-00011330>
HAL Id: tel-00011330
https://tel.archives-ouvertes.fr/tel-00011330
Submitted on 9 Jan 2006
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

Approche quantitative de la réponse écologique des
espèces végétales forestières à l'échelle de la France
Thèse de Doctorat en Sciences Forestières
de l'Ecole Nationale du Génie Rural des Eaux et des Forêts (ENGREF)
Financement : Office National des Forêts et Conseil Régional de Lorraine
Christophe Coudun
Thèse soutenue publiquement à Nancy le 22 juin 2005 devant le jury suivant:
M. Sylvain Plantureux Professeur, ENSAIA Vandoeuvre Président
Mme Sandra Lavorel Directrice de Recherche, CNRS Grenoble Rapporteur
M. Martin Hermy Professeur, Katholieke Universiteit Leuven (Belgique) Rapporteur
M. Jean-Claude Rameau Professeur, ENGREF Nancy Directeur
M. Jean-Claude Gégout Maître de Conférence, ENGREF Nancy Responsable
M. Anthony Lehmann Chargé d’étude, CSCF Neuchatel (Suisse) Examinateur
M. Patrice Mengin-Lecreulx Chef du Département Recherche, ONF Fontainebleau Examinateur
Laboratoire d’Etude des Ressources FOrêt-Bois
Unité Mixte de Recherches INRA-ENGREF 1092
VoS126
VoS125
VoS124
VoS123
2003/07/21
X : 7°21’52.04’’
Y : 48°23’22.88’’
Z : 490 m
EcoPlant

Approche quantitative de la réponse écologique des espèces végétales forestières Novembre 2005
- 2 -

Approche quantitative de la réponse écologique des espèces végétales forestières Novembre 2005
- 3 -
Remerciements
Merci à Jean-Claude Gégout pour sa gentillesse, sa disponibilité et le suivi exceptionnels qu’il
m’a accordés depuis plus de cinq ans maintenant… Les félicitations du jury sont également pour
lui !
Merci à Jean-Claude Rameau, pour son suivi et pour les nombreuses discussions matinales
autour du café. Sa disparition quelques mois après ma soutenance laissent un grand vide dans le
monde de l’écologie et de la forêt…
Merci à Christian Piedallu, pour sa disponibilité et son soutien précieux dans la manipulation
des systèmes d’information géographique, et à Ingrid Seynave, pour les calculs de valeurs
indicatrices dans le dernier chapitre !
Merci à tous les membres de l’équipe Ecologie Forestière de l’ENGREF de Nancy, qui m’ont
permis d’évoluer dans un cadre de vie et de travail remarquable, et particulièrement à Isabelle
Anselme et Evelyne Chaudy pour leur aide dans la préparation de la soutenance et du pot de
thèse, et à Damien Marage, « Pepe » Cabello et Paulina Pinto pour avoir partagé la vie de
« thésard » en même temps que moi.
Merci à Laurette Westrich, pour la reproduction de la thèse.
Merci aux nombreux auteurs des études intégrées à EcoPlant, sans qui bien évidemment mon
travail de recherche aurait été impossible.
Merci aux membres de mon comité de thèse, et particulièrement à Jean-Luc Dupouey et Jean-
Christophe Hervé pour leurs conseils écologiques ou méthodologiques avisés.
Merci aux membres du jury, et particulièrement à Sandra Lavorel et Martin Hermy pour leur
travail de rapporteur et à Anthony Lehmann, pour m’avoir permis de rencontrer mes « auteurs
préférés » lors de la semaine Riederalp en août 2004.
Merci aux membres du Centre d’Etude de la Dynamique du Carbone Terrestre (CTCD), et
particulièrement à F. Ian Woodward, Shaun Quegan et Sam Evans, pour la confiance qu’ils
m’ont accordée en m’offrant un poste trois mois avant la date de soutenance.
Merci à ma mère, mon frère et mes sœurs, pour leur soutien et pour tous les moments passés
ensemble.
Merci à mon père, parti trop tôt, pour m’avoir toujours soutenu.
Merci à Julie, pour tout le reste. Je lui dédie ma thèse…
Farnham, Angleterre, le 27 octobre 2005,
Christophe Coudun.

Approche quantitative de la réponse écologique des espèces végétales forestières Novembre 2005
- 4 -
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
1
/
129
100%