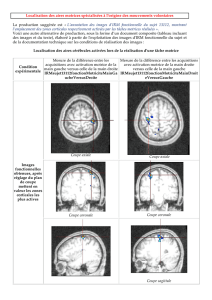
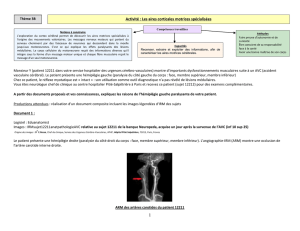
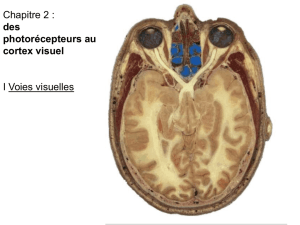
No Slide Title

Les anomalies dans la conscience de l’action
Blakemore et al. (2002)
Copie efférente: prédit les conséquences sensorielles des
commandes motrices lorsqu’on exécute des mouvements
(modèle d’anticipation)

Ataxie optique (syndrome de Balint):
difficulté (consciente) à prendre des objets
que l’on peut voir nettement
les informations visuelles associées à la forme de l’objet ne
sont pas traitées de manière correcte et donc les
commandes motrices nécessaires pour réaliser l’action sont
mal informées (mais le mouvement programmé
correspond à l’intention)

La main anarchique:
par ex., gribouillages de la main
indépendants de la volonté (dissociation
entre l’action désirée et l’action réalisée)
c’est l’opposé de l’ataxie optique, la vue d’un objet est
suffisante pour déclencher le mouvement, les effets des
indices visuels ne peuvent plus être inhibés;
mais le patient est conscient du fait que sa main ne se
conforme pas à ses intentions
«elle (la main) ne fait pas ce que je veux qu’elle fasse »

Comportement d’utilisation:
en cas de lésion frontale, utilisation stéréotypée, donc
inappropriée dans un contexte plus large, d’un objet, suite
à sa perception.
Contrairement à la main anarchique, le patient n’a pas
conscience de la dissociation entre intention et action.
Rationalisation: il l’a fait parce qu’il croyait que
l’expérimentateur voulait qu’il le fasse.
Plus précoce, dans le développement d’une action, que le
cas de la main anarchique (ne remarque l’action qu’après
l’avoir réalisée).

1. Le patient n’est pas conscient de ce qu’il fait
avant de l’avoir fait
2. Les réponses inappropriées suggérées par
les objets ne sont pas inhibées
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
1
/
36
100%