Etude des variations des traits fonctionnels de trois

Etude des variations des traits fonctionnels de trois
espèces de ligneux en réponse aux facteurs
environnementaux
(Alpes-de-Haute-Provence - 04)
BERNARD Marie, FOURRIER Camille, LADENT Emilie, POUCHARD Nicolas,
ROMERO Bastien, SCIBEK Pauline, WENDLAND Céline
M1 Master Set parcours SBE
Année 2014 / 2015

1
Résumé
L’écologie fonctionnelle des communautés vise à comprendre les mécanismes d’assemblage des
communautés, sur base des traits fonctionnels des espèces. La présence des espèces d’une
communauté est influencée par les facteurs environnementaux. Les traits fonctionnels varient selon
les contraintes abiotiques et les interactions biotiques constituant ainsi une réponse à un ou plusieurs
facteurs environnementaux. L'étude a été réalisée dans une lande à genévriers. Elle avait pour but
d'analyser les modifications des traits fonctionnels de trois espèces d’arbustes (Rosa montana, Rosa
pimpinellifolia et Berberis vulgaris) en fonction de l’altitude (de 1623m à 2100m) et de la présence de
genévrier (Juniperus communis et Juniperus sabina). Nous avons décidé de travailler sur la hauteur
végétative ainsi que le nombre de fruits.
Les résultats montrent globalement une baisse significative de la hauteur végétative et du nombre de
fruits, pour toutes les espèces sauf Rosa montana, et pour tous les milieux. Pour ce qui est de
l'influence du milieu, les traits fonctionnels sont toujours plus forts à l’extérieur des genévriers,
excepté pour Berberis vulgaris. Cette observation pourrait montrer une relation de facilitation entre le
genévrier et l'arbuste, c'est à dire que la niche écologique offerte par la plante nurse (protection contre
le froid, présence d'un humus plus épais...) permettrait à Berberis v. de mieux se développer. Ceci ne
s'observe cependant qu'à basse altitude. Pour les autres espèces, il est plus probable que la relation
inter-spécifique arbuste – genévrier soit de type neutre ou compétitrice. La compétition peut avoir lieu
au niveau de la lumière, ou au niveau des racines pour la quête de ressources nutritives.
Mots clés : traits fonctionnels, altitude, interactions biotiques, variation, écosystème montagnard
Introduction
Les recherches en écologie se sont efforcées de
mieux comprendre les processus et les patrons
de répartition des espèces en fonction des
conditions environnementales. Ainsi, depuis
une dizaine d'années, le rôle des traits
fonctionnels dans le fonctionnement des
écosystèmes fait l'objet d'un questionnement
important en écologie et motive de nombreuses
recherches.
Un trait fonctionnel est une caractéristique
morphologique, physiologique ou
phénologique ayant un impact indirect sur la
fitness de la plante via ses effets sur sa
croissance, sa reproduction et sa survie (Violle
et al. 2007). Les traits fonctionnels varient selon
les contraintes abiotiques et les interactions
biotiques (compétition, prédation, facilitation),
constituant ainsi une réponse à un ou plusieurs
facteurs environnementaux. Les traits
fonctionnels des organismes sont au cœur de
leur adaptation évolutive et écologique aux
conditions des milieux dans lesquels ils vivent.
Les conditions climatiques représentent un
premier niveau de « filtre écologique » et
déterminent ainsi le pool d’espèces susceptibles
de se maintenir dans une zone géographique
plus ou moins large (Keddy 1992). Les relations
entre organismes sont étudiées comme un
mécanisme important pour expliquer, entre
autres, l’évolution des espèces, la coexistence
des espèces, la structuration des communautés,
ou la dynamique des successions végétales
(Grassein 2011).
L’approche fonctionnelle permet donc de
faciliter l’interprétation du lien entre
composition des communautés (notamment via
les stratégies des différentes espèces
présentes) et facteurs écologiques, via
l’identification des mécanismes responsables
de cette structuration (Ansquer 2006).
Une meilleure compréhension de ces processus
permettrait d’améliorer les stratégies de
conservation ainsi que les programmes de
restauration des milieux.

2
C’est dans ce contexte que se porte notre étude,
le but étant de déterminer quelles influences
exercent les conditions environnementales sur
des espèces de ligneux dans un écosystème de
montagne.
Ce développement nous a conduits à étudier les
variations des traits fonctionnels de trois
espèces de ligneux épineux : Rosa montana,
Rosa pimpinellifolia et Berberis vulgaris selon un
facteur abiotique (l’altitude) et des interactions
biotiques dans un site situé dans les Alpes de
Hautes Provence. Nous souhaitons plus
particulièrement déterminer quel lien existe-il
entre Juniperus sabina, Juniperus communis et
les trois espèces étudiées.
Pour cela, nous nous concentrerons sur deux
traits importants : la hauteur végétative
maximale qui reflète le pouvoir de
développement de la plante et le nombre de
fruits qui détermine le pouvoir reproducteur de
la plante.
Matériels et méthodes
A. Site d’étude
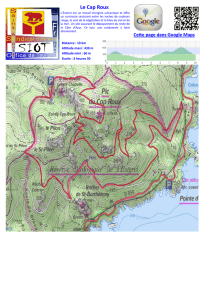
Le site d’étude se situe à Meyronnes en région
Provence-Alpes-Côte-D’azur dans les Alpes-de-
Haute-Provence (04). La zone est en versant
sud, ou adret, avec une température plus élevée
et une végétation montant plus haut qu'en
versant nord, ou ubac.
La formation végétale sous arbustive
représentative de ce site est caractérisée par
des fourrés à genévriers sabine (Juniperus
sabina). Cette formation se définit par des
zones claire et écorchée où sont présents des
affleurements rocheux et du sol nu. Elle est
principalement caractérisée par des tapis
rampants et plaqués au sol de genévrier sabine
dominant, associé à d’autres genévriers,
églantiers et arbustes adaptés aux stations
chaudes et sèches. Cette lande associe des
arbustes rampants, semi-dressés ou dressés, en
mosaïque avec des pelouses rocailleuses sèches
steppiques, éboulis et rochers (code CORINE
31.432).
Le site est prospecté par un gradient altitudinal
allant de 1623 mètres à 2110 mètres avec un
changement des conditions
environnementales. On observe ainsi une
modification graduelle des facteurs biotiques
tels que la température et l’exposition aux
rayonnements solaires mais également des
habitats et des communautés vivantes.
Ainsi le gradient altitudinal détermine
l'étagement de la végétation, avec une
communauté dominée par des genévriers
communs et sabine entre 1600m et 1900m
(étage montagnard). Passé ce palier, une forêt
de conifères est caractérisée par le pin sylvestre
avec une diminution importante de la
population de genévriers (entre l’étage
montagnard et sub-alpin). Au-delà des 2100m, il
y a une quasi absence de végétation, présence
de quelques arbustes clairsemés.
B. Récolte des données
Pour ce travail de terrain, 6 groupes ont été
établis avec l’étude de différents aspects:
caractéristique du sol, variation des traits
fonctionnels des espèces et des communautés
végétales en fonction des facteurs
environnementaux…
Le protocole d’échantillonnage s’est basé sur la
méthode d’un transect horizontal établis sur 5
stations d’altitudes différentes: 1623 m, 1830
m, 1920 m, 1995 m et 2210 m.
En ce qui concerne notre groupe d’étude, une
zone d’acceptation des individus a été établis de
chaque côté du transect, soit 10 m de part et
d’autre. Afin d’obtenir un nombre suffisant de
données, 10 individus de chaque espèces de
genévriers (commun et sabine), avec présence
d’espèces ligneuses à l’intérieur, ont été
sélectionnées, et cela pour chaque station.
Sur le versant, de nombreux tas de pierre,
nommé « clapiers », sont les vestiges d’une

3
activité agricole. Ils ont été exclus de notre
étude.
Dans un premier temps, chaque point
d’observation a été répertorié par coordonnées
GPS (Garmin GPSMAP 62s). Puis, pour chaque
groupe d’individus respectant les conditions de
notre étude, différents points ont été annoté :
L’espèce de genévrier (commun ou sabine)
Les dimensions du genévrier (longueur,
largeur, hauteur)
La ou les espèces ligneuses présentent à
l’intérieur de celui-ci
Les paramètres des traits morphologiques
potentiellement intéressants de l’espèce (la
hauteur, le diamètre, le nombre de rejets, le
nombre de fruits)
L’altitude
Afin de mesurer ces paramètres, nous avons
utilisés: un compas forestier électronique, un
décamètre, un mètre ruban et un dendromètre.
Pour les informations concernant les espèces
présentes à l’extérieur des genévriers, nous
avons eu recours la base de données d’un autre
groupe de ce travail de terrain. Et en ce qui
concerne l’aspect pédologique des stations,
nous avons également récupérés les données
d’un autre groupe. L’humus et les horizons du
sol ont été caractérisés (à l’intérieur et extérieur
des genévriers).
C. Analyses statistiques
Pour l’étude statistique, seul la hauteur et le
nombre de fruits des espèces seront analysés,
car le nombre de données est suffisant.
Les jeux de données ont été analysés avec le
logiciel R. Un test d’analyse de la variance
(ANOVA), à un facteur contrôlé a été effectué
afin de comparer les moyennes de ces
échantillons. Les P-values sont de 0.001 et 0.05.
Pour la représentation graphique, les boxplots
ont été utilisés, auxquelles sont associés la
médiane et les quartiles. Cette approche
statistique est un moyen rapide de faire figurer
le profil de la série de données.
Le modèle de régression linéaire a permis de
donner les estimations des paramètres de ce
modèle. On l’utilise avec le coefficient de
détermination R² pour voir si la variable x a un
effet sur celle de y donc ainsi remarquer s’il y a
une corrélation entre les deux variables.
Résultats
A. Rosa montana / Hauteur
L’analyse de variance (ANOVA) a montré que les
genévriers (Juniperus) et la hauteur des
genévriers ont un effet sur la hauteur de Rosa
montana.
Ces effets sont tous les deux très significatifs
avec des p-values respectives de 2e-6 et 1,37e-
8 (***). Le gradient altitudinal n’a par contre pas
d’effet sur la hauteur sur cette espèce (Fig.1).
Fig.1 : Hauteur moyenne (cm) de Rosa montana en
fonction de l'espèce de genévrier et de l'altitude (m).
Fig.2 : Hauteur moyenne (cm) de Rosa montana en
fonction de l'espèce de genévrier.

4
La médiane de la hauteur du Rosa montana au
sein du genévrier commun est de 50 cm, et de
160 cm dans le genévrier sabine de 160 cm, elle
est plus importante à l'extérieur des genévriers,
qui est égale à 200 cm (Fig.2).
On observe une corrélation négative entre la
hauteur des genévriers et la hauteur de Rosa
montana dont la régression linéaire possède un
R² ajusté de 0,36. Ceci montre la forte influence
de la taille du genévrier hôte sur la hauteur de
Rosa montana. Plus il est grand, moins le Rosa
montana est développé.
B. Rosa montana / Nombre de fruits
L’analyse de variance (ANOVA) a montré que les
genévriers (Juniperus) (Fig.3) et la hauteur des
genévriers ont un effet sur le nombre de fruits
de Rosa montana. Le nombre de fruits est
équivalent entre les espèces de genévriers mais
est significativement plus important à
l'extérieur (p-value= 6,45e-5 ***).
Fig.3: Nombre de fruits chez Rosa montana en fonction de
l'espèce de genévrier.
Il existe une corrélation négative entre la
hauteur des genévriers et le nombre de fruits,
celle-ci est exprimée par un R² ajusté de 0,265.
C. Rosa pimpinellifolia / Hauteur
L’analyse de variance (ANOVA) a montré que les
genévriers (Juniperus) et l’altitude ont un effet
sur la hauteur de Rosa pimpinellifolia. Ces effets
sont tous les deux très significatifs avec des p-
values respectives de 6e-13 et de 2e-16 (***).
L’effet de la hauteur des genévriers est moins
significative (*).
Il est important de noter l’absence surprenante
de représentation des Rosa p. le long du
gradient altitudinal. Ce biais méthodologique
sera repris dans la discussion.
On observe une baisse de la hauteur des Rosa p.
le long du gradient altitudinal. La régression
linéaire associée à un R² de 0.49 (Fig.4), ce qui
indique que la moitié de la variance de la
hauteur est corrélée à ce facteur.
Fig.4: Régression linéaire entre la hauteur (cm) de Rosa p.
et le gradient altitudinal (m).
D. Rosa pimpinellifolia / Nombre de fruits
L’analyse de variance (ANOVA) a montré que les
genévriers (Juniperus), l’altitude et la hauteur
des genévriers ont un effet sur les fruits de Rosa
pimpinellifolia. Les effets des genévriers et de
l’altitude sont tous les deux très significatifs
(***). L’effet de la hauteur des genévriers est
moins significative (*).
Fig.5 : Régression linéaire entre le nombre de fruits chez
Rosa p. et l'altitude (m).
 6
7
8
9
6
7
8
9
1
/
9
100%