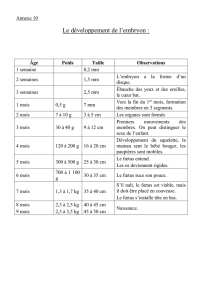
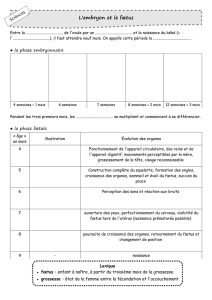
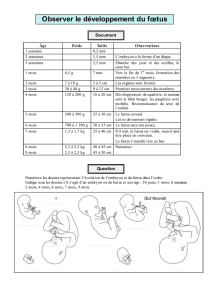
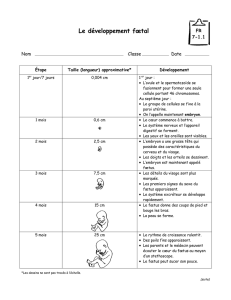
ÉTUDE BIBLIOGRAPHIQUE DE L`UTILISATION DU FŒTUS DE

ÉCOLE NATIONALE VÉTÉRINAIRE D’ALFORT
Année 2012
ÉTUDE BIBLIOGRAPHIQUE DE
L’UTILISATION DU FŒTUS DE BREBIS
COMME MODÈLE EXPÉRIMENTAL
THÈSE
Pour le
DOCTORAT VÉTÉRINAIRE
Présentée et soutenue publiquement devant
LA FACULTÉ DE MÉDECINE DE CRÉTEIL
par
Marie-Aude, Annette, Christiane GENAIN
Née le 7 Avril 1987 à Paris 15ème
JURY
Président : Pr.
Professeur à la Faculté de Médecine de CRÉTEIL
Membres
Directeur : M. Christophe DEGUEURCE
Professeur à l’ENVA
Co-Directeur : M. Luc Behr
Directeur Scientifique et technique à IMM Recherche
Assesseur : M. Renaud TISSIER
Maître de Conférences à l’ENVA




REMERCIEMENTS
Au président du jury, Professeur à la faculté de Médecine de Créteil,
Qui nous fait l’honneur de présider notre jury,
Hommage respectueux.
A Monsieur Christophe Degueurce,
Professeur à l’Ecole Nationale Vétérinaire d’Alfort, Conservateur au Musée Fragonard
Qui a accepté d’encadrer ce travail de thèse,
Grâce à qui j’ai pu participer à la vie du Musée, et découvrir l’histoire de son siècle racontée avec tant de passion,
Amitiés et sincères remerciements.
A Monsieur Luc Behr,
Directeur Scientifique et technique à l’IMM Recherche
Qui m’a proposé ce sujet passionnant et a accepté de diriger ce travail,
Pour sa disponibilité, le temps qu’il m’a consacré, son enseignement, ses conseils, sa sympathie et nos échanges,
You’re simply the best!
A Monsieur Renaud Tissier,
Maître de conférences à l’Ecole Nationale Vétérinaire d’Alfort,
Qui nous a fait l’honneur d’accepter de participer à notre jury de thèse,
Sincères remerciements.
A toute l’équipe d’IMM Recherche, qui m’a si chaleureusement accueillie, et avec qui c’est toujours un immense
plaisir de travailler. Par ordre alphabétique de prénoms, à : Alain, Anta, Christophe, Hervé, Jean-Philippe, Lamia, Luc,
Marie-Pierre, Mehrak, Nicolas, Rémi, Sébastien et Virginie. Pour tout ce que chacun d’entre vous m’a apporté comme
enseignement, soutien, conseils, bonne humeur, fous rires, humanité, sympathie, et j’en oublie. MERCI !!!!!!!
A Monsieur Yves Millemann, Maître de conférences à l’ENVA, sans qui l’aventure à l’IMM n’aurait jamais pu
débuter. Pour son esprit d’ouverture, son écoute et ses conseils. Merci.
To all the outstanding and inspiring specialists that I was very fortunate to work with and who made my externships
not only possible, but memorable. For their warm welcome, and their precious help and advice : Drs D. Begon, L
Behr, N. Borenstein, B. Biller, P. Boscan, P. Early, R. Hess, B. Keene, C. MacPhail, W. Maï, E. Monnet, B. Posch, D.
Smeak, and S. Withrow. And to each one of their residents and interns, who taught me so much !
Aux enseignants d’Alfort, à qui je dois l’apprentissage de ce métier-passion et une extraordinaire formation humaine.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
1
/
136
100%