31/10/2013 HAMEL Audrenne L2 G. KAPLANSKI 14 pages

TISSU SANGUIN ET SYSTEME IMMUNITAIRE - BASES GENERALES
Inflammation
31/10/2013
HAMEL Audrenne L2
Tissu sanguin et système immunitaire-bases générales
G. KAPLANSKI
14 pages
Titre du cours
INTRODUCTION
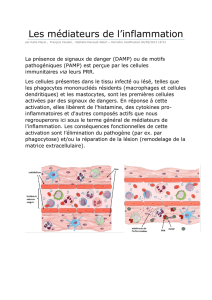
La réaction inflammatoire (RI) peut-être considérée comme la réponse immunitaire innée. Mais elle est bien
différente le la Réponse Immunitaire adaptative.
Définition : c'est un mécanisme de défense essentiel à l’organisme pour lutter contre les agressions tissulaires
de nature variables, c'est à dire microbiennes ou blessures d'un tissu.
Le but de cette réaction est de circonscrire l'agent agresseur dans son milieu, et de le détruire pour revenir au
niveau de base, l'homéostasie.
Les mécanismes impliqués sont : la margination, l’extravasation (sortie des vaisseaux sanguins), et la
migration des leucocytes sanguins.
Souvent, les leucocytes font aussi des dommages collatéraux dans les tissus. Afin d'être bénéfique, la RI doit
donc être limitée dans l’espace et dans le temps : c'est la RI aiguë.
1/14
Plan
PARTIE 1 : la réaction inflammatoire aiguë
A. La phase vasculaire
B. La phase cellulaire
I. les cellules impliquées
1. leucocytes
a) Mastocytes
b)polynucléaires neutrophiles
c) monocytes et macrophages résidents
2. les cellules endothéliales
3. les fibroblastes
II. Médiateurs solubles
III. Médiateurs membranaires : molécules d'adhésion
IV. Cascade adhésive leuco-endothéliale
V. Migration trans-endothéliale
VI.Passage à travers la MEC
C. La phase résolutive
I. Fin de la Réaction inflammatoire
II. Comment induit-on l'inflammation ?
PARTIE 2 : La réaction inflammatoire chronique
I. Dérégulation du système inflammatoire : maladie auto-inflammatoire
II. Persistance de l'agent agresseur
III. Auto-immunité
IV. Dérégulation due à un état anormal chronique

TISSU SANGUIN ET SYSTEME IMMUNITAIRE - BASES GENERALES
Inflammation
Si elle n'est pas limitée, on a affaire à une RI chronique ou diffuse : on qualifie la maladie d’inflammatoire
(c'est le cas de très nombreuse maladies actuelles).
La RI aiguë est un mécanisme essentiel à notre survie : elle est plus précoce que l’immunité adaptative et
prépare le terrain pour cette dernière. Mais contrairement à elle, la RI aiguë ne possède pas de mémoire : elle
se met en place au bout du 2 ou 3ième contact avec l'agent agresseur.
L'inflammation peut être induite par deux sortes de mécanismes :
•une attaque microbienne
•elle peut aussi survenir en milieu stérile comme pour :
•la crise de goutte causée par une accumulation d'acide urique dans une articulation
•la Silicose : exposition à la silice qui pénètre dans les poumons
•la nécrose cellulaire qui est sans doute responsable de la plupart des inflammations en
déclenchant le système immunitaire.
PARTIE 1 : LA REACTION INFLAMMATOIRE AIGUE
Elle se compose de 3 phases:
–la phase vasculaire , qui dure quelques minutes ou quelques heures. On assiste à une vasodilatation, à
une augmentation de la perméabilité des capillaires et de la coagulation, ainsi qu'à une fuite de plasma (œdème)
–la phase cellulaire, qui dure quelques heures à quelques jours. On assiste à une cascade d'adhésion
leucocytes-cellules endothéliales, à la migration trans-endothéliale et tissulaire des leucocytes ainsi qu’à la
destruction de l'agent agresseur.
–La phase résolutive , qui dure plusieurs jours. C'est la fin de la RI. On assiste à une reconstitution du
tissu lésé, notamment par la production de facteurs de croissance.
Où se passe cette RI ?
Au niveau des capillaires et post-veinules car le flux sanguin y est beaucoup plus lent que dans les autres
vaisseaux de diamètre plus élevé. Il peut y avoir une interaction leucocytes-paroi des cellules endothéliales. De
plus, les conditions de perméabilité de cette zone facilitent le passage des leucocytes hors du sang.
A. La phase vasculaire
Elle se produit très rapidement, au bout de quelques secondes et dure quelques heures.
(ex : un pneumocoque qui entre dans les poumons va déclencher cette phase en quelques secondes).
Elle résulte de l'action de nombreux médiateurs protéiques ou lipidiques vaso-actifs ayant une réaction rapide
sur le tonus vasculaire et préformés ou stockés et disponibles immédiatement.
Il y a des médiateurs préformés et stockés tels que : Histamine et Sérotonine.
Ou encore des protéines plasmatiques inactives, s’activant rapidement : Hageman (facteur XII) ou le
Complément (C3a,C5a,C4a).
On trouve aussi des médiateurs lipidiques qui ne seront synthétisés et sécrétés qu'au bout de quelques
minutes : les dérivés de l'acide arachidonique tels que les Prostaglandines E2 et 12, les leucotriènes, etc..
2/14

TISSU SANGUIN ET SYSTEME IMMUNITAIRE - BASES GENERALES
Inflammation
Le tonus vasculaire va diminuer, entraînant une vasodilatation, donc une diminution du flux sanguin. Cela
entraîne une augmentation de la perméabilité de la membrane endothéliale et donc un efflux de protéines
plasmatiques : il y a création d'un œdème.
B. La phase cellulaire
I. les cellules impliquées
1. les leucocytes
a) les Mastocytes
Ce sont les cellules résidentes sentinelles car elles sont au contact du milieu extérieur. On les trouve dans le
tube digestif, dans les bronches, sphère ORL,...elles peuvent donc recevoir les informations concernant le
danger.
Sur leur membrane cellulaire, on trouve un grand nombre de récepteurs : FcεR, FcγR, complément, Toll,
cytokines,...
Ces cellules possèdent une capacité de réponse rapide et brève qui leur permet de participer à la phase
vasculaire. En effet, elles contiennent des molécules préformées comme des eicosanoides (dérivés de l'acide
arachidonique), de l'histamine, de la sérotonine...
Elles ont aussi une capacité de réponse plus lente et plus prolongée par la production de cytokines et
chimiokines ce qui permet la participation à la phase cellulaire.
b) les Polynucléaires neutrophiles (PnN)
Ce sont les principales cellules effectrices de la RI.
Elles ont la capacité d'infiltrer les tissus grâce à l'expression de molécules adhésives telles que les L-sélectines,
ICAM, β2 Intégrine..), grâce à des récepteurs à des signaux chimiotactiques (leucotriènes, PAF-Facteur
d'Activation Plaquettaire, chimiokines..) qui les attirent sur le site de l'inflammation, ainsi que grâce à leur
appareil locomoteur (pseudopodes...)
3/14

TISSU SANGUIN ET SYSTEME IMMUNITAIRE - BASES GENERALES
Inflammation
Les PnN ont une capacité de phagocytose, de bactéricidie et de lyse. Ils ont des récepteurs opsonines, fabriquent
des radicaux libres et dérivés de l'O2. Les enzymes qui permettent la destruction des agents microbiens sont par
exemple les protéinases 3, les élastases, cathepsine...
opsonine : fragment qui se trouve sur la membrane des bactéries. les Pn possèdent des récepteurs à ces
fragments, ce qui leur permet de les reconnaître et de les détruire.
c) Les monocytes et macrophages résidents
Ces cellules possèdent une capacité d'infiltration tissulaire et de destruction : ils ont des molécules adhésives
(ICAM , β1 et β2 Intégrine), des récepteurs pour la synthèse de chimiokines (mobilité), des récepteurs pour la
phagocytose, des enzymes (myéloperoxidase), des radicaux libres...
Par la production de cytokines et des chimiokines, ces cellules participent à l'amplification de la Réaction
Inflammatoire (RI).
Elles participent aussi à la résolution de la RI en phagocytant les neutrophiles apoptotiques (qui ont une vie
courte). Comme les macrophages et les monocytes présentent le complexe HLA II (=CMH II) à leur surface, ils
peuvent induire la réponse immunitaire adaptative qui se fera par production d'anticorps.
2. les cellules endothéliales
Elles contrôlent le tonus vasculaire (qui est modifié par la vasodilatation), mais aussi l'hémostase (coagulation),
la cascade d'adhésion des leucocytes aux cellules endothéliales et leur migration (mise en jeu des molécules
suivantes : PECCAM, ICAM-I, VCAM-I, E-sélectine, cadhérine, production de chimiokines).
3. les fibroblastes
Ils migrent vers le site inflammatoire, où ils ont une fonction essentielle de réparation des tissus lésés. Ils
produisent alors du collagène, de la Matrice Extra-Cellulaire (MEC), des enzymes, des facteurs de croissance...
II. Médiateurs solubles
1) Cytokines
Ce sont des protéines de petit PM (10-20kDa) qui sont sécrétées par différentes cellules en réponse à une
agression de type inflammatoire, avec activation via les récepteurs Toll-like. Elles transmettent l'information de
l'inflammation aux autres cellules, on les considère comme les hormones de l'inflammation.
Certaines sont sécrétées très précocement (1à 2h après le début de l’agression): Interleukines (IL) ou TNF
(Tumor Necrosis Factor).
D'autres plus tardivement, avec effet limité : les chimiokines.
TNF et IL-1 ont une action pléiotrope : elles agissent sur différents types cellulaires et permettent différents
types d'action.
4/14

TISSU SANGUIN ET SYSTEME IMMUNITAIRE - BASES GENERALES
Inflammation
a) IL-1
L'interleukine 1 fait partie de la famille des cytokines (IL-1a, IL-1b, IL 18, IL 33 ou encore IL-1-Ra qui est un
antagoniste du récepteur à IL-1a)
Sur cette diapo, nous avons une comparaison des gènes de différentes IL : IL-1a et IL-1b.
La * signifie que la molécule est déjà active : le précurseur de l'IL-1a (PM 31kDa) est actif tandis que celui de
l'IL-1b ne l'est pas et nécessitera un clivage par une caspase 1 pour atteindre sa forme mature de 17kDa.
On notera toutefois que le précurseur d'IL-1a est clivé par une calpaine pour atteindre un PM de 17kDa. Cette
forme mature est également active.
Au niveau des récepteurs des ces IL, il en existe 2 types : un actif, IL-1R type 1 qui est activé par la fixation
d'IL sur lui et un autre récepteur, IL-1R type 2 qui lui n'est pas actif : il sert de piège à IL.
En effet, une dose de 100 picogrammes/mL de cytokines entraînerait la mort. L'organisme doit donc toujours en
contrôler les doses.
IL-1 est synthétisée par toutes les cellules, et surtout par les macrophages et les monocytes.
IL-1a n'est pas sécrétée, elle est cytoplasmique. Par contre si la membrane cellulaire devient poreuse comme
lors d'une nécrose, alors on la retrouve en dehors de la cellule (signal d'une inflammation).
IL-1b est sécrétée de manière soluble.
b) TNF α
Le précurseur du TNF α est une molécule transmembranaire active de 31 kDa qui est active biologiquement
lorsque deux cellules entrent en contact.
Pour sa maturation, ce précurseur est clivé par TACE (TNF Converting Enzyme) en une molécule soluble de
17kDa. Les deux récepteurs qui lui sont associés sont actifs.
5/14
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%