Texte intégral View

UNIVERSITÉ DE NEUCHÂTEL INSTITUT DE ZOOLOGIE
Modulation de l'immunité de lapins contre les
tiques Ixodes ricinus L.: effets de la charge
parasitaire et de traitements par les cytokines
IL-2 et TNF-ct
par
Sandra Schorderet
Licenciée en Biologie
Thèse présentée à la Faculté des Sciences de l'Université de Neuchâtel pour
l'obtention du grade de docteur es sciences.
1993

à Jean-Marc
à mes parents

IMPRIMATUR POUR LA THÈSE
Modulation de 1'immunité de lapins contre
les
..tiques.
Ixodes ricinus..
L...:
elf eta..de
Ia charge parasitaire et de traitements
par""ie&""eytöklriöB'"iE-2'"'et""TRP-a
de Mademoiselle...Sandra...Schorderet
UNIVERSITÉ DE NEUCHÂTEL
FACULTÉ DES SCIENCES
La Faculté des sciences de l'Université de Neuchâtel
sur le rapport des membres du jury,
MM.
M. Brossard, B. Rutti, K. Pfister
(Berne) et S. Martinod (Lincoln, Nebraska)
autorise l'impression de la présente thèse.
Neuchâtel,
le ^...août 1993
Le doyen :
A. Robert

TABLE DES MATIÈRES i
TABLE PES MATIÈRES
1.
Introduction 1
1.1. Présentation du parasite 1
1.2. Les hôtes développent une résistance contre les tiques 1
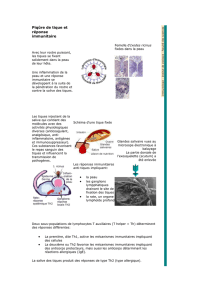
1.3. La peau de l'hôte, organe-cible des tiques 2
1.4. La piqûre des tiques provoque une inflammation locale
de la peau de l'hôte , 4
1.5. La piqûre des tiques induit chez l'hôte une réponse immunologique
spécifique 7
1.6. Les tiques peuvent prévenir les défenses de l'hôte 10
1.7. Caractérisations d'antigènes 12
1.8. Les cytokines, un nouvel outil pour comprendre et moduler
la réponse immunologique d'un hôte face à un parasite 13
1.8.1. IL-2 13
1.8.2.
Le TNF-a, médiateur des réactions inflammatoires 14
1.9. Buts du travail : 18
2.
Matériel et méthodes 19
2.1.
Animaux 19
2.1.1.
Tiques 19
2.1.2. Lapins 19
2.2.
Antigènes 19
2.2.1.
Extrait de glandes salivaires (EGS) 19
2.2.2. Extrait tégumentaire (ET) 20
2.2.3.
Récolte de salive 20
2.3.
Effets de la charge parasitaire sur l'acquisition de la résistance
et la réponse immunitaire cellulaire et humorale du lapin 21
2.3.1.
Expérience préliminaire 21
2.3.2. Infestations en continu 21
2.3-3.
Modulation de la réponse cellulaire et humorale du lapin
par deux degrés d'infestaticn différents, et conséquences
sur le développement de la résistance 21
2.4. Effets d'un traitement à ITL-2 recombinante humaine
sur l'acquisition de la résistance chez des lapins infestés,
et sur leur réponse cellulaire et humorale contre les tiques 22
2.4.1.
Antigène 22
2.4.2. IL-2 22
2.4.3.
Première expérience 22
2.4.4. Expérience complémentaire 23
2.5.
Effets du TNF-a sur l'acquisition de la résistance et sur
la réponse immunitaire des lapins infestés 23
2.5.1.
Antigène 23
2.5.2. TNF-a 23
2.5.3.
Vaccination contre le TNF-a 23
2.5.4. Traitement des lapins au TNF-a 24
2.6.
Effets in vitro des antigènes de tiques sur les lymphocytes et les
neutrophils de lapins indemnes ou sensibilisés par les tiques 24
2.6.1.
Effet sur les lymphocytes 24
2.6.2. Effet sur les neutrophiles 25

TABLE DES MATIÈRES H
2.7.
Effets du sérum de lapins sensibilisés aux tiques sur la
prolifération de lymphocytes d'animaux indemnes ou sensibilisés 25
2.8.
Statistique 25
3.
Résultats et discussions 26
3.1.
Effets de la charge parasitaire sur l'acquisition de la résistance
et la réponse immunitaire cellulaire et humorale du lapin 26
3.1.1.
Expérience préliminaire 26
3.1.1.1.
Remarques générales 26
3.1.1.2. Biologie des tiques 26
3.1.1.3.
Sérologie 28 •
3.1.2. Infestations en continu 28
3.1.2.1.
Remarques générales 28
3.1.2.2. Biologie des tiques 29
3.1.2.3.
Sérologie 29
3.1.3.
Commentaire intermédiaire '. 31
3.1.4. Modulation de la réponse cellulaire et humorale du lapin par
deux degrés d'infestation différents, et conséquences sur le
développement de la résistance 31
3.1.4.1.
Biologie des tiques 31
3.1.4.2. Réponse cellulaire des lapins:
analyse par transformation lymphoblastique 32
3.1.4.3.
Sérologie 33
3.1.5.
Discussion 33
3.1.6. Conclusion 37
3.2. Effets d'un traitement à 1TL-2 recombinante humaine sur
!'acquisition de la résistance chez des lapins infestés,
et sur leur réponse cellulaire et humorale contre les tiques 37
3.2.1.
Remarque générale 37
3.2.2. Première expérience 38
3.2.2.1.
Effet du traitement à I'IL-2 sur la biologie
des tiques 38
3.2.2.2. Sérologie 38
3.2.2.3.
Transformation lymphoblastique 39
3.2.3.
Considérations intermédiaires 40
3.2.4. Expérience complémentaire 40
3.2.4.1.
Remarque préliminaire 40
3.2.4.2.' Réponse cellulaire des lapins 40
3.2.4.3.
Biologie des tiques 42
3.2.5.
Discussion 43
3.2.6. Conclusion 46
3.3.
Effets du TNF-ct sur l'acquisition de la résistance et sur
la réponse immunitaire des lapins infestés 47
3.3.1.
Remarque préliminaire 47
3.3.2. Vaccination contre le TNF-a 47
3.3.2.1.
Titres d'anticorps anti-TNF-o 47
3.3.2.2. Réponse humorale contre I1EGS 47
3.3.2.3.
C3 sérique 48
3.3.2.4. Réponse cellulaire des lapins:
analyse par transformation lymphoblastique 48
3.3.2.5. Biologie des tiques 49
3.3.3.
Remarques 49
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
1
/
142
100%