11coursTSI.7

http://lewebpedagogique.com/bouchaud
1
I.7 Immunologie.
I.7 Immunologie.
Introduction.
Le système immunitaire regroupe l’ensemble des cellules d’un individu pouvant s’opposer à la
pénétration et à l’infection par des agents pathogènes.
Les cellules du système immunitaire sont les leucocytes (= globules blancs). Ils comprennent
notamment les lymphocytes B (LB) sécréteurs d’anticorps, les lymphocytes T (LT) détruisant les cellules
infectées ou mutées, les phagocytes (ex : monocytes dans le sang et macrophages dans les tissus) effectuant la
phagocytose. Ces cellules sont produites dans la moelle osseuse rouge (organe lymphoïde primaire, comme le
thymus).
Ces cellules se regroupent dans les organes lymphoïdes secondaires (ganglions lymphatiques, rate…)
ou dans le sang.
À partir de l’exemple du SIDA, on va voir comment le système immunitaire intervient lors d’une
infection puis, dans un deuxième temps, sur quelle propriété du système immunitaire reposent les vaccins.
1. Comment se manifeste le SIDA ?
Le SIDA, ou syndrome (= ensemble des manifestations de la maladie) d’immunodéficience acquise, est
une maladie d’origine virale, conduisant à la destruction progressive du système immunitaire.
Cette maladie est planétaire : on parle de pandémie.
1.1 Le virus VIH.
Un virus est une particule de petite taille, constituée d’un acide nucléique (ADN ou ARN) et d’une
enveloppe protéique parfois doublée d’une enveloppe lipidique. Le virus doit se servir des enzymes et des
organites de la cellule infectée pour se multiplier : c’est un parasite intracellulaire obligatoire.
Le SIDA est dû au virus VIH (virus de l’immunodéficience humaine) dont il existe deux souches :
VIH-1 et VIH-2.
Le virus VIH.
© http://artic.ac-besancon.fr/svt/act_ped/svt_lyc/eva_bac/s-bac2008/images/vih.gif
Ce virus de 100 à 120 nanomètres de diamètre possède :
- Une enveloppe lipidique contenant des protéines, notamment
gp120 et gp41, qui constituent des antigènes.
- Deux capsides, constituées de p17 et p24 (une seule
représentée sur l’image) renfermant 2 ARN et des molécules
d’enzymes (transcriptase inverse). Comme c’est un virus à
ARN, on parle de rétrovirus.
Les organes lymphoïdes.
© http://pst.chez-
alice.fr/svtiufm/images/sysimmun.gif
- Organes lymphoïdes centraux =
primaires. Ce sont les lieux de
production et de différenciation des
lymphocytes. A noter que les LT
deviennent matures dans le thymus.
- Organes lymphoïdes périphériques =
secondaires.

http://lewebpedagogique.com/bouchaud
2
Un antigène est une molécule étrangère à un organisme pouvant déclencher une réaction immunitaire.
Ce virus est transmis par voie sexuelle, par voie sanguine ou au cours de la grossesse de la mère à
l’enfant (particulièrement à l’accouchement ou par allaitement).
Les cellules cibles du VIH sont principalement les cellules immunitaires : les lymphocytes T4 (voir
2.3), les monocytes et les macrophages (voir 2.1).
Un macrophage est cellule immunitaire provenant de la transformation d’un monocyte (cellule immunitaire
circulant dans le sang) dans les tissus. Les deux ont pour fonction la phagocytose.
Les LT4, les monocytes et les macrophages sont les cibles du virus car ils possèdent des protéines
membranaires (CD4) auxquelles le virus s’amarre par l’intermédiaire d’une protéine de son enveloppe (gp120).
Cette fixation lui permet de pénétrer dans la cellule hôte.
Une fois dans la cellule, le virus doit se multiplier.
Une enzyme virale, la transcriptase inverse, rétro-transcrit l’ARN viral en ADN dans les cellules
infectées. Cet ADN est intégré au génome de la cellule et s’exprime, permettant la reproduction du virus sous
forme de particules virales infectieuses et leur dissémination dans l’organisme. Voir le cycle du virus.
Une fois dans l’organisme, le virus atteint les organes lymphoïdes (ganglions lymphatiques), où sont
localisées les cellules immunitaires. Dans ces organes lymphoïdes, les lymphocytes T4 et les macrophages
présents sont infectés et jouent ainsi le rôle de réservoir à virus.
Particules de virus du SIDA (VIH)
bourgeonnant à la surface d'un
lymphocyte T4 infecté. Vue au
microscope électronique, après
coloration.
© http://www.cnrs.fr/cnrs-
images/sciencesdelavieaulycee/org_animal/popup_
immuno5.htm

http://lewebpedagogique.com/bouchaud
3
LE CYCLE DU VIH
Cellules cibles : LT4, macrophages, monocytes. Il faut entre 1 ou 2 jours à 1 ou 2 semaines (suivant l’état de la
cellule cible) pour que le virus accomplisse son cycle.
© Hatier TS 2002, modifié 04/05
Ancrage du virus à la cellule cible
(reconnaissance gp120 – CD4)
GP120
CD4
Fusion des membranes et pénétration de la
capside contenant les ARN viraux.
Libération des ARN viraux
Rétrotranscription des ARN
viraux en ADN, puis intégration
dans le génome cellulaire.
Transcriptase
inverse
Production d’ARNm viraux par
transcription grâce à la machinerie
cellulaire.
Traduction en protéines virales
Assemblage des virus
Bourgeonnement de nouveaux virus
GP120
Transcriptase
inverse

http://lewebpedagogique.com/bouchaud
4
On distingue plusieurs étapes à partir de la contamination.
1.2 Étape 1 : la primo-infection.
Pendant cette période de quelques semaines, les symptômes se limitent le plus souvent à ceux d’une
maladie virale bénigne (grippe légère). Les virus se multiplient rapidement causant une forte baisse du nombre
de LT4. Les anticorps anti-VIH apparaissent et neutralisent les virus présents dans la circulation. De même, les
LTc anti-VIH apparaissent et détruisent les cellules infectées (monocytes, macrophages et LT4). La conséquence
est la remontée du nombre de LT4.
Un lymphocyte T cytotoxique provient de la différenciation des lymphocytes T8, et est responsable de la
destruction des cellules infectées, mutées ou greffées.
La présence d’anticorps anti-VIH deux semaines à quelques mois après la contamination montre que le sujet
est « séropositif » pour le VIH. Ces anticorps sont spécifiques contre certaines protéines du VIH (gp120,
gp41…). Ce sont ces anticorps qui sont décelés par le test ELISA (Enzyme Linked Immuno Sorbent Assay) et le
Western-blot (pour confirmer un test ELISA positif) (bien entendu, il faut environ au moins 20 jours post-
contamination pour les détecter d’où des ELISA qui peuvent être négatifs).
1.3 Étape 2 : la phase asymptomatique.
Comme son nom l’indique, c’est la phase sans symptôme : le sujet se porte bien pendant plusieurs
années, en moyenne 10 (durée très variable d’un individu à l’autre). Mais on constate toutefois :
- un affaiblissement progressif du nombre de LT4.
- une faible charge virale.
- une forte présence d’anticorps anti-VIH et de LTc anti-VIH, preuve que le virus est toujours présent.
En fait, les défenses immunitaires luttent activement, mais les virus continuent à se multiplier dans les
LT4, macrophages et monocytes (réservoirs). En conséquence, les lymphocytes T4 infectés sont détruits par les
LTc, et malgré leur renouvellement, leur nombre diminue.
1.4 Étape 3 : la phase symptomatique (stade SIDA déclaré).
En absence de traitement, la charge virale augmente fortement et le nombre des LT4 baisse. Lorsqu’il
est suffisamment bas (< à 200 par mm3 de sang), diverses maladies opportunistes apparaissent. Ce sont des
maladies dont le développement est habituellement contrôlé par le système immunitaire. Ces maladies
surviennent du fait de l’affaiblissement des défenses de l’organisme.
Les LT4 ont donc un rôle majeur dans les processus immunitaires.
Résultat de TP du test ELISA.
©
http://lewebpedagogique.com/bouchaud/files/Photo0077.
jpg
Exemple de test Western Blot.
© http://artic.ac-
besancon.fr/svt/act_ped/svt_lyc/eva_bac/s-
bac2009/images/western-blot.gif

http://lewebpedagogique.com/bouchaud
5
PRIMO
INFECTION
PHASE ASYMPTOMATIQUE
SIDA
* fièvre
* pas de signes cliniques
* maladies
opportunistes
* réponse
immunitaire
initiale
* réponse immunitaire qui limite
le développement du virus
* mauvais
fonctionnement
du système
immunitaire
* déficience
immunitaire
totale
Les différentes phases du SIDA. © Didier TS 2002, modifié 04/05
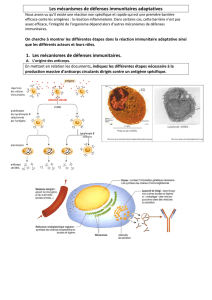
2. Comment le système immunitaire intervient-il lors d’une infection ?
Les mécanismes immunitaires se répartissent en deux ensembles :
- L’immunité innée, assurée par les phagocytes. Très rapide. Image : « médecin généraliste ».
- L’immunité acquise correspond à l’intervention des LB et LT suite au premier contact avec l’antigène
(plus lent). Image : « médecin spécialiste »
2.1 Les anticorps sont les agents du maintien de l’intégrité du milieu extracellulaire.
Un anticorps est une protéine produite par les plasmocytes, en réponse à la présence d’un antigène et capable
de le reconnaître spécifiquement.
- Ce sont les antigènes eux-mêmes qui déclenchent leur prolifération (ils existent déjà même si l’AG n’a jamais
été rencontré).
La molécule d’anticorps sous le
logiciel RasTop.
©
http://www.inrp.fr/Acces/biotic/rastop/exemp
les/images/bilan.jpg
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%