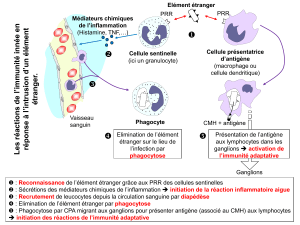
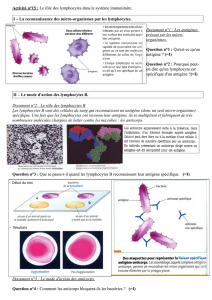
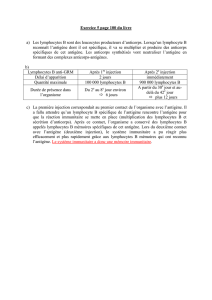
le récepteur spécifique du lymphocyte t - iPubli

Armand
Bensussan
à
Violaine
David
ADRESSE
-
L
RÉFÉRENCEs
2,

C
14
s
D
4
• 297
1
/
2
100%