HBDD 4 CHAPITRE 1 (2ème partie) 3) Développement du

HBDD 4 CHAPITRE 1 (2ème partie)
3) Développement du processus chordal
D18
*Petite vidéo sur le DVD*
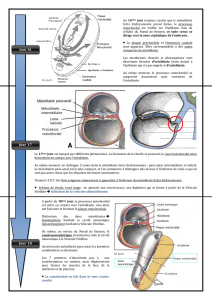
Cette vidéo va être consacrée à la mise en place du processus chordal. Vous avez à gauche
une coupe sagittale où vous reconnaissez toujours la cavité amniotique à la face dorsale de
l’embryon, la cavité vitelline à la face ventrale, l’endoderme en jaune, l’ectoderme en bleu
et le mésoblaste en rouge et le processus chordal en vert. Dans la partie droite des schémas
vous avez une coupe transversale dont le plan de coupe vous est figuré sur la coupe
sagittale. Vous reconnaissez l’ensemble des structures déjà dénommées.
On se place dans un premier temps en coupe transversale vous retrouvez la migration des
cellules du mésoblaste pour retrouver le processus chordal, ou notochordal process. C’est
un processus canalaire, une invagination en doigt de gant, borgne à son extrémité crâniale.
Vous le voyez sur la coupe sagittale en haut, et vous voyez une structure tubulaire sur la
coupe transversale en bas. Le nœud de Hensen est dénommé ici primitive pit. Dans un
premier temps, le plancher de ce processus chordal va s’ouvrir à la face ventrale de
l’embryon et les cellules vont devenir jointives avec les cellules de l’endoderme ce qui
deviendra la plaque chordale ou notochordal plate. Cette plaque chordale en vue sagittale à
gauche et transversale à droite. La ligne pointillée correspond toujours au plan de section
correspondant à la vue transversale. Cette ouverture du plancher pour former
temporairement une plaque chordale va permettre la communication par l’intermédiaire
du canal neurentérique entre la vésicule vitelline à la face ventrale et la cavité amniotique à
la face dorsale de l’embryon.
Dans un deuxième temps ce processus voit son plancher se refermer pour former à
nouveau un processus tubulaire qui cette fois sera plein (le schéma est un peu faux) et sera
dissocié de l’endoblaste sous-jacent. On parle alors de chorde définitive ou notochorde.
Vous voyez donc sur cet embryon au niveau du mésoblaste la visualisation de la notochorde
sous forme de ligne verte qui s’arrête en crânial juste avant la membrane bucco pharyngée.
La zone entre la membrane bucco pharyngée et le processus chordale sera la plaque
préchordale. Le reste du mésoblaste est en rouge.

4) Régionalisation du mésoblaste
D19.
Vous avez donc vu dans la précédente vidéo la mise
en place du mésoblaste dans ses différentes
portions : Le processus chordal qui va aboutir à la
formation de la chorde définitive.
Evolution spécifique des différentes régions du
mésoblaste selon l’axe médio-latéral :
--- Axial : processus chordal qui deviendra la chorde définitive
--- Para---axial : Lieu de la mise en place de la
somitogénèse.
La différenciation du mésoblaste donnera ici la
future structure vertébrale de l’embryon.
--- Intermédiaire : (non étudié dans ce cours) formation du
système urogénital
--- Latéral : Creusé d’une cavité qui permettra la
formation de 2 feuillets : splanchnopleure et
somatopleure
5) Organisation du mésoblaste para-axial
a. Formation des somitomères
D20.
Voyons dans un premier temps le mésoblaste para axial, avec la formation des
somitomères et ultimement la formation des somites. Tous ces évènements sont
dynamiques.
La différenciation progressive des cellules du mésoblaste para axial pour former des
somitomères se fait donc au fur et à mesure que le processus chordal se met en place.
Cette différenciation se fait dans le sens crânio-caudal elle va donc débuter dans la zone
médiane de l’embryon et progresser vers la partie crâniale de l’embryon.
On aura donc les structures les plus différenciées en position plus crânial, c’est un gradient
de différenciation.

Cette diapo illustre l’embryon en coupe transversale, vous reconnaissez l’ectoblaste en
bleu, avec la zone de la plaque neurale en bleu. On retrouve la partie ventrale
l’hypoblaste, et le mésoblaste en rouge.
En bas de la diapo on voit une photo d’un embryon en coupe transversale en microscopie
électronique, on retrouve ectoblaste, processus chordal, mésoblaste para axial,
intermédiaire et latéral.
On voit les différentes zones du mésoblaste en rouge, le reste en bleu et jaune. La zone en
orange correspond à des zones de décollement, mais n’existe pas in vivo, en réalité les
feuillets sont ben accolés.
: Embryon en coupe transversale (représentation schématique en haut,
microscopie électronique à balayage en bas).

b. Transformation des somitomères en somites
D21.
Afin de mieux comprendre la mise en place du mésoblaste para axial en sens crânio-caudal,
on a sur cette diapo un embryon en 3D coupé transversalement. Dans la partie haute on a
l’organisation dorso ventrale de l’embryon, t l’organisation médiane et latérale du
mésoblaste. Vous repérez la partie dorsale en bleu (ectoblaste) et la partie médiane en
rouge correspondant au mésoblaste.
Dans la partie basse on peut visualiser la chorde et le mésoblaste para axiale vue en
microscopie électronique à balayage. Cet embryon est vu cette fois par sa face dorsale, et
on a retiré tout le feuillet ectoblastique afin de mieux visualiser le mésoblaste.
En 4 la chorde ou notochorde, et en 1 le mésoblaste para axiale, qui commence à se former
en boule, comme un collier de perle qui se met progressivement en place.
Cette individualisation de zones sous forme de boules successives correspond au passage
du mésoblaste para axial aux somitomères puis somites. La mise en place de ces
somitomères débutera dans la partie moyenne de l’embryon, au niveau du nœud de
Hensen pour la ligne primitive. La progression est ensuite de la partie crâniale à la partie
caudale.
On voit sur la figure ci-dessous le processus chordal orienté dans l’axe crânio-caudal au
niveau de la partie médiane de l’embryon.
: Schéma et photographie en microscopie électronique à balayage d’un embryon en vue
dorsale. Sur la microphotographie, le feuillet ectoblastique a été enlevé.

c. Somitogenèse
: Schématisation d’un embryon au niveau médian et para---axial dans le sens longitudinal.
La mise en place des somites se fait dans le sens crânio--- caudal, on trouve donc au
niveau crânial les stades les plus avancés et au niveau caudal les stades les moins avancés
de la somitogenèse.
Le premier stade de formation des somites se fait à partir des cellules mésenchymateuses
du mésoblaste.
Ces cellules ont une organisation de type tissu conjonctif lâche.
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%