La réponse immunitaire Il existe 3 types de réponse immunitaire qui

La réponse immunitaire
Il existe 3 types de réponse immunitaire qui vont résulter de la coopération entre les différentes
cellules:
- réponse immunitaire qui conduit à la formation des LTCD4 helper
- réponse immunitaire qui donne naissance, à partir des lymphocytes B, aux plasmocytes (→
formation d'anticorps)
- réponse immunitaire qui aboutit aux phénomènes cytotoxiques
La réponse immunitaire est déclenchée par la rencontre avec l'antigène.
Antigène = molécule susceptible d'être reconnue par le BCR ou le TCR:
- qui stimule la production d'une réponse immunitaire
- qui réagit avec les produits de la réponse immunitaire: les anticorps
Un antigène va comporter de nombreux épitopes.
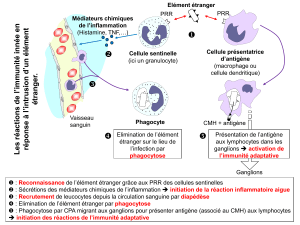
I) Coopération entre les LTCD4 et une cellule présentant l'antigène
Ca conduit à la formation des LTCD4 helper.
Diapo 2+3
Un lymphocyte B reconnaît une partie de l'antigène, l’antigène n'a pas besoin d'être modifié
(reconnaissance directe).
Un lymphocyte T reconnaît un fragment de l'antigène (peptide) sur la membrane d'une autre cellule
(présentation de l'antigène).
A) Cellules présentatrices d'antigènes (CPAg)
Toute cellule présentant des molécules de classe II du CMH est potentiellement une CPAg pour les
lymphocytes T CD4.
- lymphocytes B
- macrophages
- cellules dendritiques
Cellules dendritiques (progéniteurs CD34+):
Ce sont les cellules les plus importantes pour la présentation des antigènes. Ces cellules sont
d'origine hématopoïétique. Il existe 2 catégories de cellules dendritiques:
- origine myéloïde: cDc (CD11c/CD18), présentent l'antigènes aux LTCD4
→ cellules dendritiques conventionnelles
- origine lymphoïde: CD plasmacytoïde (pDC). Produisent de l'IFN si activées et ne
présentent pas « bien » l'antigène aux LTCD4
→ rôle dans la défense anti-virale
cDC: lorsqu'elles sortent de la moelle, on dit qu'elles sont dans un état immature. Elles vont aller
dans les tissus (sous les épithélia, organes solides comme le rein, le cœur, le foie, …). Dans la peau,
elles prennent le nom des cellules de Langerhans.
Les pDC sont essentiellement retrouvés dans les organes lymphoïdes.
Diapo 4
Quand elles arrivent dans les tissus périphériques, les cellules dendritiques sont totalement
immatures. Elle est capable d'endocyter, de phagocyter. Elles n'expriment quasiment pas de

molécules de classe II du CMH, mais elles expriment des molécules de classe I. Elles n'expriment
pas de molécules de co-stimulation (CD40, CD80, CD86).
Diapo 5
En cas d'inflammation ou d'infection, ces cellules sont attirées par des chémokines et elles vont être
activées par les ligands du TLR (LPS, ARNdb, …), des cytokines, … A ce moment là, ces cellules
vont migrer par voie lymphatique pour aller dans les ganglions lymphatiques proches, changer
l'expression de certains récepteurs, etc. Au cours de cette migration vers les organes lymphoïdes,
elles vont perdre leur capacité d'endocytose et de phagocytose, elles vont exprimer des molécules de
classe II du CMH en réponse à l'activation du TLR, et elles vont exprimer des molécules de co-
stimulation.
Quand elles arrivent dans les ganglions, elles sont maintenant capables de présenter l'antigène
qu'elles ont phagocyté et de stimuler les lymphocytes T.
Diapo 6
D'autres cellules sont capables de réaliser la présentation de l'antigène aux lymphocytes T CD4:
• Macrophages:
- capables de phagocyter, d'endocyter, de macropinocyter)
- capables d'exprimer des molécules de costimulation
- tissus lymphoïdes et conjonctifs, cavités corporelles
- l'expression des molécules de classe II du CMH est constitutive ou inductible par des
cytokines ou des bactéries
• Lymphocytes B:
- présents dans les tissus lymphoïdes
- expression des molécules de classe II du CMH inductible au cours de la maturation
- les molécules de costimulation sont inductibles au cours de la maturation
Les cellules dendritiques présentent l'antigène à des lymphocytes T naïfs, alors que les macrophages
et les lymphocytes B présentent l'antigène à des lymphocytes T effecteurs.
Les cellules dendritiques, les macrophages et les lymphocytes B ont besoin d'être activés pour
exprimer les molécules de costimulation, les molécules de classe II du CMH, … Les antigènes du
soi ne sont pas capables de stimuler cette expression.
B) Interactions cellulaires
La réponse immunitaire comporte 5 étapes:
- reconnaissance par le lymphocyte T
- activation
- prolifération
- génération des effecteurs
- régulation
La reconnaissance de l'antigène va dépendre de sa voie d'introduction:
• Si l'antigène est introduit par voie cutanée, la réponse se fait en général dans les ganglions
régionaux. Cet antigène va être transporté par la lymphe ou par les cellules dendritiques.
• Si l'antigène pénètre par voie sanguine, la réponse se fait dans la rate.
• S'il pénètre au niveau des muqueuses, la réponse se fera par le système lymphoïde associé aux
muqueuses et par les ganglions voisins.

1) Phase de reconnaissance
Un lymphocyte T CD4 ne reconnaît un antigène que s'il est présenté par des molécules de classe II
du CMH. Ces molécules de classe II du CMH n'insèrent que des peptides dans leur cavité, donc un
lymphocyte T CD4 classique ne reconnaît qu'un antigène de nature protéique.
Cet antigène est internalisé, dégradé dans des endosomes, puis les molécules de classe II du CMH
vont associer ces peptides et les présenter à leur surface.
La reconnaissance d'un antigène par un LTCD4 va dépendre de son internalisation, de sa
dégradation, de son association avec les molécules de classe II du CMH et surtout de la
reconnaissance par le TCR.
2) Etape d'activation
Diapo
Les cellules vont entrer en contact les unes avec les autres. Une fois que la reconnaissance est
établie, il va y avoir une transduction de signaux, puis la transcription de gènes (≈ 100) dans le
lymphocyte T. Ils vont également exprimer de nouvelles molécules à la fois solubles et
membranaires et ils vont entrer en prolifération.
Quand ces lymphocytes T sont dans les ganglions, ils vont établir des contacts avec les cellules
dendritiques, grâce à CD2 (ligand = LFA3) et ICAM (ligand = LFA-1). Ces contacts sont
transitoires mais si le TCR reconnaît un peptide, les lymphocytes T s'arrêtent à la surface des
cellules dendritiques. L'interaction renforcée se fait par LFA-1 – ICAM1. Il faut qu'il voit, à la
surface de la cellule dendritique, au moins 10 à 50 peptides pour être activé.
Il va se former, entre le lymphocyte T et la cellule dendritique, une synapse immune. Au centre de
cette synapse se trouvent le TCR, CD2/LFA3, … et sont repoussées en périphérie des molécules
dont la partie extracellulaire est beaucoup plus longue (phosphatases qui pourraient interrompre le
signal).
Diapo
Cette synapse se produit au niveau de radeaux lipidiques (raft), où se trouvent les molécules qui
vont permettre la transmission du signal.
Diapo
Transmission du signal:
La première étape est le recrutement de tyrosine-kinases de la famille src: lck, fyn. Au moment de la
formation de la synapse, ces 2 kinases sont rapprochées et activées, et elles vont phosphoryler les
motifs ITAM les chaînes zéta des CD3. Ces motifs phosphorylés vont recruter une autre tyrosine-
kinase appelée ZAP-70, qui a des motifs SH2 (lui permettent d'interagir avec les motifs ITAM
phosphorylés). ZAP-70 est phosphorylée par lck et fyn.
→ 1ère étape = recrutement et phosphorylation de ZAP-70
ZAP-70 va ensuite phosphoryler 2 molécules adaptatrices:
- LAT
- SLP-76
Diapo
Ca permet le recrutement et l'activation de:

• La phospholipase Cγ (PLCγ) qui va hydrolyser le PIP2 en:
- DAG qui active les PKC → activation de NF-κB
- IP3 qui augmente le Ca2+ intracellulaire, ce qui active la calcineurine → activation de NF-
AT
• Activation de la voie de ras et des protéines G de la famille Rho, ce qui conduit à l'activation de la
voie des MAPkinases → activation d'AP-1.
La plupart des immunosuppresseurs agissent au niveau de la voie NF-AT (bloquent l'effet de la
calcineurine).
Une fois que ce système est activé, il faudra des co-signaux qui permettent:
- la survie
- la prolifération
Il va y avoir expression de CD40L à la surface du lymphocyte T. Son interaction avec CD40 permet
à la cellule présentant l'antigène une augmentation de l'expression des molécules de co-stimulation.
Ces molécules (B7) vont interagir avec le CD28 (= signal 2), ce qui va stabiliser le messager d'une
cytokine qui est l'interleukine-2. S'il n'y a pas ce signal, il n'y a pas de prolifération. Le signal 2 va
également activer la voie de la PI3kinase.
Le troisième signal indispensable est fourni par les cytokines produites par les CPAg.
L'activation conduit à la transcription de nombreux gènes:
• Transcription immédiate: protooncogènes cellulaires: c-fos, c-jun, c-myc (quelques minutes)
Ces molécules sont impliquées dans la croissance cellulaire et dans la prolifération.
• Transcription précoce: gènes de cytokines, dont l'IL-2 (cette cytokine permet la suite!)
Facteurs de transcription du gène de l'IL-2:
- SP-1 et Egr-1 (early growth response 1 protein)
- NF-AT
- AP-1
- OCT-1
- cREL (famille des facteurs de transcription REL)
- NF-κB (protéines p65 et p50)
- HMG-I(Y) ou high mobility group of proteins
Il y a également transcription de récepteurs de cytokines, dont le gène qui code pour la chaîne α du
récepteur de l'IL-2. Il y aura ainsi apparition d'un récepteur à haute affinité de l'IL-2.
• Transcription tardive: HLA-DR, VLA-1, 2, 3, 4, 5 (15 jours)
Ces intégrines vont permettre aux lymphocytes d'aller ailleurs que dans les organes lymphoïdes.
3) La prolifération
La prolifération va apparaître au bout de 48 à 72 heures. L'IL-2 a un rôle autocrine. Tous les
immunosuppresseurs utilisés actuellement ont pour cible l'IL-2 (production ou interaction avec le
récepteur).
L'IL-2 va induire la prolifération des lymphocytes (multiplication 2 à 3 fois par jour, pendant
plusieurs jours). On va obtenir des clones de lymphocytes T CD4, qui ont tous le même TCR. Ces
clones vont avoir des propriétés différentes. On parle de LTCD4 helper (ou auxiliaire).
On a 3 types de LTCD4 helper qui vont apparaître:
- LTh1
- LTh2
- LTh17
Tout ceci dépend de la différenciation des cytokines produites par les cellules dendritiques
présentant l'antigène.

• Les LTh1:
La cellule dendritique va produire de l'interféron-γ qui va activer un facteur de transcription (STAT
1) qui induit un autre facteur de transcription (T-bet). On voit apparaître, sur le lymphocyte T, le
récepteur de l'IL-12, ce qui active STAT 4 (facteur de transcription). Ces facteurs de transcription
vont transformer le lymphocyte T CD4 naïf en un LTh1. Ces facteurs de transcription vont permettre
à cette cellule de produire de l'interféron-γ. Les LTh1 sont responsables de l'immunité cellulaire:
il aide d'autres cellules à se débarrasser de tout ce qui est pathogène intracellulaire, grâce à
l'interféron-γ.
Ils activent les lymphocytes T CD8, les cellules NK, les macrophages.
Ces cellules jouent un rôle défavorable dans les maladies auto-immunes. Les LTh1 disparaissent lors
d'une infection par HIV. Ils aident les lymphocytes B à produire les Ig G.
• Les LTh2:
Ici, la cellule dendritique ne produit pas d'interféron-γ, mais, sous l'influence de bactéries
extracellulaires, de parasites ou d'allergènes, il y a production d'IL-4. Il y a 2 facteurs de
transcription importants: STAT6 et NF-AT. Le GATA 3 intervient également. Le LT naïf va se
transformer en LTh2, qui va produire d'autres cytokines dont l'IL-4.
Les LTh2 sont responsables de l'immunité humorale. Ils vont aider les lymphocytes B à produire
des anticorps (IgM, IgE, IgA). Les anticorps ne peuvent être actifs que sur tous les pathogènes
extracellulaires (bactéries, parasites, …).
• Les LTh17:
Le TGF-β et L'IL-6 activent RORγT, ce qui induit la transformation des LT naïfs en LTh17. Ils
produisent l'IL-17, ce qui active les macrophages et les polynucléaires neutrophiles.
Il faut absolument que ce système soit régulé!
La réponse Th1 bloque la réponse Th2 et la réponse Th2 bloque la réponse Th1. La meilleure
régulation est réalisée par les lymphocytes T régulateurs, qui vont arrêter la réponse immunitaire.
Il existe des LT régulateurs naturels, produits dans le thymus, et des LT régulateurs induits en
périphérie.
Ils agissent par des contacts cellule-cellule et surtout en produisant des cytokines inhibitrices dont
l'IL-10.
Parmi les cellules produites vont apparaître des cellules mémoires (LT mémoire). Ces cellules ont
une durée de vie très longue. On les retrouve dans les organes lymphoïdes et dans les tissus. Ces
cellules sont activables très facilement. La présence de ces lymphocytes T mémoires à vie très
longue explique la vaccination et le maintien du nombre constant de LT alors que le thymus involue
après la puberté.
1
/
5
100%