Dans la science contemporaine, il n`y a qu`une théorie de l`évolution

Gène. Séquence de nucléotides formant une unité d’information génétique.
Allèle. Chacune des différentes séquences (ou versions) d’un même gène.
1. La diversité (ou variation) génétique : les mutations
http://www.snv.jussieu.fr/vie/dossiers/evolution/evol/traits_2.html
Mutation. Modification aléatoire de la séquence des nucléotides de l’ADN.
Au sens strict une mutation est due à des erreurs lors de la réplication ou à des agents externes (agents mutagènes)
et ne touche donc qu’une ou quelques bases. De manière plus générale on appelle aussi mutation toute modification
héréditaire du matériel génétique due à des éléments mobiles, des rétrovirus, des recombinaison, des duplications
ou des transpositions. Dans ce cas la modification peut toucher plusieurs dizaines ou centaines de nucléotides, voire
de chromosomes entiers.
L’impact d’une mutation est très variable. S’agissant d’un phénomène aléatoire il est beaucoup
plus fréquent d'introduire dans un gène une modification qui en supprimera ou en modifiera la
fonction qu'une mutation qui l’améliore. Cependant le nombre de divisions cellulaires menant aux
gamètes est faible par rapport au nombre conduisant aux cellules somatiques. Il en résulte que le
nombre d'erreurs de copie est également limité (mais non nul). Nous nous intéresserons seulement
aux mutations qui touchent la lignée germinale, puisque ce sont les seules qui peuvent être
transmises à la descendance.
D’autres facteurs peuvent atténuer l’impact réel d’une mutation.
- La position de la mutation sur le gène peut n’avoir aucune conséquence (mutation synonyme) ou modifier
de manière plus ou moins importante totalement la protéine codée (mutations faux sens et non sens).
- Plus de 90% du génome n'a pas de fonction connue, et une grande partie. Les modifications se produisant
dans ces régions ajoutent à la variabilité, mais n'ont pas d'influence sur le phénotype et la valeur sélective.
- Dans un certain nombre de cas, il suffit d’une seule copie fonctionnelle du gène pour que sa fonction
assurée, partiellement ou dans son entier. Les hétérozygotes porteurs d’un allèle avec une modification qui
serait théoriquement létale peuvent n’être pas ou très peu affectés et seuls les homozygotes seront contre-
sélectionnés.
- Il existe des protéines chaperons dont le rôle est d'assurer l'acquisition de la bonne conformation d'autres
protéines même si la séquence de celles ci est modifiée.
- De nombreuses voies métaboliques comprennent des redondances, de sorte qu’une inactivation complète
de la protéine codée par un gène peut ne provoquer aucun changement notable dans le fonctionnement de
l'organisme touché.
- Lorsqu'un gène a été dupliqué, la modification même radicale de l'une des copies ne provoquera aucun
désagrément à l'organisme, puisqu’il existe une deuxième copie fonctionnelle.
Une proportion importante des mutations et modifications sont neutres (pas d’effet sur la valeur
sélective des individus). Cela explique une certaine stabilité de l’espèce au cours du temps. Les
mutations qui « parviennent » à passer et à avoir des effets constitueront le « carburant » de
l’évolution.

2. Le tri sur la diversité : dérive génétique et sélection naturelle
http://www.snv.jussieu.fr/vie/dossiers/evolution/evol/tridiv_1.html
Si la sélection s'exerce sur le phénotype au niveau individuel, la compréhension de la variation
des caractères au cours du temps et donc des processus évolutifs demande que l'on s'intéresse à une
échelle plus large : des ensembles d'individus ou populations.
Une population est un ensemble d’individus qui ont la même probabilité de se croiser entre eux et qui se
reproduisent moins, ou pas du tout, avec les membres d’autres populations. Cette reproduction diminuée est due à
des critères extrinsèques, spatiaux par exemple, et non à des barrières génétiques entre les groupes ; les populations
ne sont pas des espèces. Cependant, il peut y avoir une différenciation génétique entre populations, avec des
fréquences alléliques, voire des allèles spécifiques, pour chacune (comme par exemple dans le cas des groupes
sanguins dans les populations humaines).
La diversité génétique résultant des mutations fait qu'à un instant donné, une population n'est
pas homogène d'un point de vue génétique : il existe pour la plupart des gènes plusieurs allèles
dont les fréquences (fréquences alléliques) vont varier au cours du temps. Un des sujets d'intérêt
particulier est l'étude de la manière dont une mutation sur une séquence (donc présente chez un seul
individu et sur une seule de ses deux copies du même locus), peut se répandre dans une population.
Cela fait intervenir deux mécanismes : la sélection naturelle et la dérive génétique.
La sélection naturelle permet la survie et/ou confère un avantage reproductif aux individus qui
se distinguent des autres par au moins un caractère héritable. La pression de sélection peut être liée
à l’influence du milieu ou à de une compétition intra (capacité à se reproduire…) ou interspécifique
(prédation, accès à la nourriture…). Ce qui est important, c'est non seulement le taux de survie,
mais également le nombre de descendants laissé.
Un variant a beau être extrêmement avantageux, s'il ne transmet pas la variation à la génération suivante, il
n'aura pas d'impact sur l'évolution à long terme de sa population ou de son espèce. Inversement, des allèles
produisant un effet négatif important ne seront pas toujours. Par exemple, un allèle létal, à partir du moment où il
est récessif, peut rester présent un temps dans une population mais ne pas se fixer, puisqu'il est létal à l'état
homozygote. De même, un allèle dominant, mais dont l'effet négatif se fait sentir après que les individus se soient
reproduits, peut être maintenu (chorée de Huntington). D’autre part, les mécanismes de crossing-over, dont la
fréquence est dépendante de la distance entre les loci, peuvent intervenir. Ainsi, si un allèle profite d’une sélection
positive, les allèles d’autres gènes situés à faible distance de celui-ci peuvent être « entraînés » par cette sélection et
fixés également, même s’ils sont neutres ou légèrement délétères.
Si la sélection naturelle tend à diminuer la fréquence des allèles désavantageux, de nombreux
allèles ne sont ni avantageux, ni désavantageux et donc ne sont pas ou peu soumis à la sélection
(allèles neutres : par exemple les allèles résultant de mutation synonymes). Le devenir de ces allèles
ne dépend que du taux de mutation (néomutations) et de la dérive génétique.
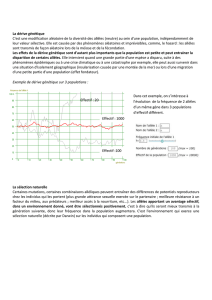
Dérive génétique est la modification, au sein d'une population, de la fréquence d'un allèle d'une
génération à l'autre. Elle est indépendante des mutations, de la sélection naturelle et des migrations.
Elle est seulement liée à des phénomènes aléatoires et imprévisibles comme, par exemple, le hasard
des rencontres des gamètes. Si une population possède un grand nombre d’individus la fréquence
des allèles neutres reste stable d’une génération à l’autre. Dans le cas d’une population de faible
effectif, les croisements se faisant au hasard, la distribution de deux allèles dans les descendants
peut être différente de celle dans la génération parentale de sorte qu’un allèle peut disparaître.
L’allèle restant est alors dit fixé et tous les descendants en seront porteurs. et tous les descendants
en seront porteurs. Si n est le nombre d’exemplaires d’un nouvel allèle dans une population, le
temps qu’il met pour envahir la population est en moyenne de 2n générations. Ainsi, plus la taille de
la population est réduite, plus un allèle peut se fixer rapidement.
La plupart des nouveaux allèles seront éliminés par ces processus il en résulte une diminution de
la biodiversité. Cependant, tôt ou tard, de nouvelles mutations se produisent, créant ainsi de
nouveaux allèles, et les mécanismes se répètent.

Voir http://www.snv.jussieu.fr/vie/dossiers/evolution/evol/derive.html
Cette explication à l'aide de deux allèles est bien sûr schématique. De nouveaux allèles au même locus peuvent
apparaître avant que ceux qui étaient précédemment présents dans la population ne soient fixés ou perdus. Ainsi, à
un instant donné, dans une population, il existe souvent plusieurs allèles pour un même locus
Si n’existe aucun processus d'homogénéisation entre les deux populations (échange de migrants
pour assurer des échanges d'allèles), après un certain temps les différences seront suffisantes pour
que les deux populations ne soient plus compatibles et interféconds c’est la spéciation
Les importances relatives de la sélection, de la dérive génétique et de la vitesse de mutation le
sont fruit de nombreux événements contingents. Les modifications du génome se font de manière
aléatoire, et non en fonction de l'avantage ou du désavantage que cela procurera à l'organisme.
Aucune évolution biologique n’est donc prévisible.
3. L’apparition d’une nouvelle espèce : la spéciation
http://www.snv.jussieu.fr/vie/dossiers/evolution/evol/tridiv_4.html
Une espèce est une population, ou un ensemble de populations, dont les individus peuvent
effectivement ou potentiellement se reproduire entre eux et engendrer une descendance viable et
féconde, dans des conditions naturelles.
Ainsi, l'espèce est la plus grande unité de population au sein de laquelle le flux génétique est possible alors que
les individus d'une même espèce sont génétiquement isolés d’autres ensembles équivalents du point de vue
reproductif.
Cependant le critère d’interfécondité ne peut pas toujours être vérifié : c'est le cas pour les fossiles, les
organismes asexués ou pour des espèces rares ou difficiles à observer. D’autres définitions peuvent donc être
utilisées :
- espèce morphologique : groupe d'individus défini par des caractéristiques structurales (taille, forme…) ;
- espèce phylogénétique : la plus petite lignée d’une population pouvant être définie par une combinaison
unique de caractères diagnostiques ;
- espèce écologique : groupe d’organismes partageant une même niche écologique ;
- espèce phénétique : ensemble d’organismes vivants se ressemblant plus entre eux qu’à d’autres ensembles
équivalents.
Il existe au moins deux types de spéciation.
La spéciation allopatrique est la plus courante. Elle se produit quand deux populations de la
même espèce sont isolées dans l’espace :
- soit l’aire de répartition est coupée en deux par l’apparition d’un obstacle ;
- soit une partie de la population s’isole par migration (ex. colonisation d’une île).
La spéciation sympatrique se fait par apparition d’une nouvelle espèce sur une aire de
répartition chevauchante avec celle de l’espèce d’origine. La séparation des deux populations est
alors liée à un isolement génétique (ex. ploïdie chez certains végétaux…) ou comportemental (ex.
période de reproduction, comportement reproducteur, préférences sexuelles…).
Quelle que soit l’origine de l’isolement les deux populations subissent alors de manière
indépendant les mécanismes de mutation, de dérive génétique et de sélection. Il en résulte
l’apparition d’une incompatibilité génétique, physiologique ou comportementale qui empêche les
individus d’un groupe de se reproduire avec les individus de l’autre groupe, même s’ils sont remis
en contact.
4. La disparition d’une espèce : l’extinction
http://www.snv.jussieu.fr/vie/dossiers/evolution/evol/tridiv_5.html
L’extinction est la disparition complète d’une espèce ou d’un groupe d’espèces. Une espèce est
considérée comme éteinte lorsque le dernier individu meurt.

D’une manière générale, l’extinction se produit lorsqu’une espèce ne peut plus survivre ou se
reproduire dans un milieu, et est incapable de gagner un nouveau milieu qui lui conviendrait.
Elles peuvent être dues par exemple à des changements brutaux du milieu (c’est le cas de
nombreuses espèces de grenouilles, de nos jours), de la prédation (on peut évoquer le cas du dodo),
ou de la compétition (certaines espèces introduites se multiplie rapidement, au détriment d’espèces
locales : grenouille taureau, caulerpe…). Des maladies peuvent également être impliquées (la grippe
aviaire qui a récemment causé la mort de plusieurs tigres vivant en liberté), ainsi que des causes
génétiques, qui frappent particulièrement les petites populations (consanguinité et/ou la dérive
peuvent être responsables de la fixation de mutations délétères). Enfin, l’extinction peut également
être due à la disparition ou extinction d’une ressource dont dépendait l’espèce pour sa survie ou sa
reproduction : pollinisateur, proie… Bien entendu, il est également possible que plusieurs facteurs
se combinent…
1
/
4
100%