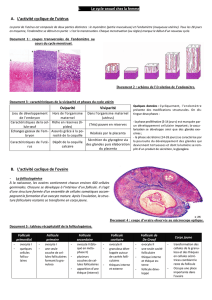
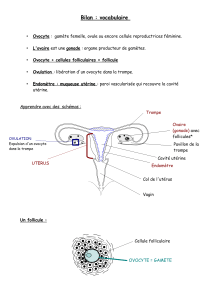
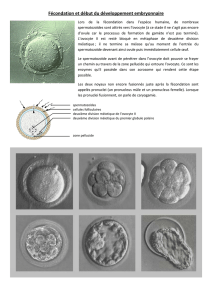
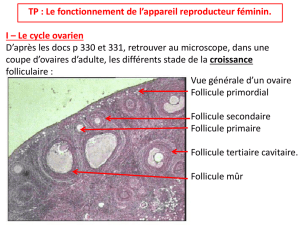
CHAPITRE I LES GAMETES 1.1. Définition Chez les espèces

CHAPITRE I LES GAMETES
1.1. Définition
Chez les espèces diploïdes, les gamètes sont des cellules haploïdes spécialisées dans la
fusion sexuée. Elles sont spécifiques d'espèce. De plus, nos animaux domestiques sont
qualifiés d'hétérogamétiques (par opposition aux espèces isogamétiques) car les deux sexes
produisent des gamètes morphologiquement distincts :
- l'un est volumineux, immobile et rare : l'ovule provenant de la femelle.
- l'un est petit, mobile et abondant : le spermatozoïde provenant du mâle.
Leur formation relève des mêmes mécanismes fondamentaux dont le principal est la
réduction chromatique ou méiose par laquelle le nombre de chromosomes est réduit de moitié,
passant ainsi de l'état diploïde à l'état haploïde.
1.2. Origine et formation : La gamétogenèse
Chez les deux sexes, les gamètes proviennent des cellules germinales primordiales
(CGP) diploïdes. On les détecte d'abord à l'extérieur du corps de l'embryon, au sein du
mésoderme extraembryonnaire sous-jacent au feuillet endodermique du sac vitellin, tout près
de l'origine de l'allantoïde en formation (figure 1.1). L'origine exacte de ces cellules
germinales primordiales est inconnue, mais certaines de leurs caractéristiques plaident en
faveur d'une origine ectodermique. Elles dériveraient ainsi de l'ectoderme embryonnaire et
migreraient jusqu'au mésoderme vitellin où elles initient leur activité phosphatase alcaline
rendant possible leur détection. Par la suite, elles poursuivent leur migration jusqu'aux
ébauches des gonades : les crêtes génitales.
Chez les oiseaux, cette migration se fait par voie sanguine. Les cellules germinales
primordiales migrent dans le système vasculaire et, arrivées dans les capillaires des crêtes
génitales, elles passent activement au travers de l'endothélium vasculaire pour coloniser la
gonade en formation (figure 1.2). Chez les mammifères, les cellules germinales primordiales
migrent par mouvements amoeboïdes en passant dans l'épaisseur de l'intestin primitif
(archentéron) pour remonter par le mésentère dorsal et gagner les crêtes génitales situées à la
face médiale des corps de Wolff (figure 1.1). Dès leur entrée dans les crêtes génitales, elles
prennent le nom de gonies. Là, elles exercent sur les tissus gonadiques une action inductive
qui oriente la différenciation de l'organe en testicule ou en ovaire selon le sexe génétique. A
ce stade, elles prennent le nom de spermatogonies ou d'oogonies (ou ovogonies).
Chez les deux sexes, la gamétogenèse passe par trois phases :
- Phase de multiplication
Les oogonies et les spermatogonies se multiplient par mitoses successives, conservant
donc leur caractère diploïde. Cette phase de multiplication, initiée durant la migration, se
poursuit activement pendant une période plus ou moins étendue suivant l'espèce (tableau
1.1). Elle se termine par l'entrée en méiose des oogonies chez la femelle et, simultanément,
par le ralentissement voire l'arrêt complet des mitoses des spermatogonies chez le mâle
(phénomène de quiescence observé chez les rongeurs).

- Phase d'accroissement - prophase méiotique
Les gonies de dernière génération cessent de se diviser et augmentent de volume, se
transformant en "cytes" de premier ordre ou "cytes I". L'augmentation de volume est très
faible dans le cas du spermatocyte I mais considérable chez l'ovocyte I car elle correspond à
l'accumulation des réserves cytoplasmiques nécessaires aux premières divisions de
l'embryon.
A ce stade, les cytes I sont encore des cellules diploïdes. Leur ADN est constitué de deux
compléments, un paternel et un maternel, eux-mêmes constitués d'une série identique de n chromosomes (n étant
un nombre caractéristique de l'espèce, voir tableau 1.2). Chaque chromosome est constitué d'un filament
d'ADN. Une cellule diploïde possède donc 2n chromosomes répartis en (n-1) paires de chromosomes
homologues ou autosomes et une paire de chromosomes hétérologues ou sexuels.
Les cytes I de dernière génération répliquent d'abord leur ADN durant la phase S de leur cycle. Ils
deviennent donc tétraploïdes. On suppose que chaque chromosome reste accolé au chromosome néo-synthétisé
correspondant formant une anse chromosomiale. Le cyte I possède donc 4n chromosomes répartis en 2n anses
chromosomiales. Les cytes I s'engagent ensuite dans la première phase de la méiose : la prophase I (figure 1.3).
Chaque anse chromosomiale. s'individualise (stades pré-leptotène et leptotène) et s'apparie avec l'anse
homologue dérivée de l'autre complément au niveau de son centromère mais également en de nombreux autres
points d'ancrage pour former les complexes synaptonémaux (stade zygotène). Le nucléole disparaît. Les anses
se condensent, s'épaississent (stade pachytène) et individualisent leurs deux chromatides-soeurs. Chaque couple
de chromatides-soeurs correspond à un chromosome et à sa copie néo-synthétisée. A ce stade, la cellule possède
4n chromosomes répartis en n tétrades (figure 1.4). Les couples de chromatides-sœurs se séparent de leur
homologue (stade diplotène) mais restent accolées par les points d'ancrage (ou sites de recombinaison) où
s'opèrent des échanges de gènes paternels et maternels (chiasmatypie ou crossing-over).
- Phase de maturation
Au cours de cette phase, les cellules germinales subissent successivement deux
divisions de maturation (voir figures 1.3). En fin de maturation, chaque cellule contient n
chromosomes et est donc authentiquement haploïde. La réduction chromatique est achevée.
Première division de maturation (DM1) : le cyte I se divise en 2 cytes de deuxième
ordre (cyte II). La membrane nucléaire disparaît et le premier fuseau achromatique se met en
place. Les tétrades se positionnent à l'équateur (stade métaphase I). Les couples de
chromatides-sœurs se séparent de leurs homologues pour se répartir équitablement et au
hasard entre les deux cellules-filles (stade anaphase I). Le cytoplasme du cyte I se divise
(cytodiérèse) pour individualiser les deux cellules filles (stade télophase I). Chaque cyte II
hérite donc de 2n chromosomes répartis en n couples de chromatides-sœurs (ou dyades). Ces
cytes II sont donc diploïdes.
Interphase : c'est la période séparant les deux divisions de maturation. Elle est très
courte. L'ADN se décondense partiellement et le centromère de chaque dyade se dédouble.
Deuxième division de maturation (DM2) : le cyte II se divise en deux "ides". Cette
division succède à l'interphase (figure 1.3). Les dyades se recondensent (stade prophase II), le
second fuseau achromatique se met en place et les dyades migrent à l'équateur (stade
métaphase II). Les chromatides-sœurs se séparent et se distribuent équitablement entre les
deux cellules filles (stade anaphase II). Le cytoplasme du cyte II se divise (cytodiérèse) pour
individualiser les deux ides (stade télophase II).

Des erreurs, d'autant plus nombreuses que les cellules germinales sont plus vieilles,
peuvent se produire au cours de la distribution des chromosomes entre les cellules filles,
entraînant des aberrations chromosomiques (trisomies, monosomies,.....). Ces aberrations,
sources de malformations fœtales graves, sont le plus souvent létales.
Si les mécanismes fondamentaux de la gamétogenèse sont identiques dans les deux
sexes, il existe de profondes différences chronologiques et morphologiques entre l'oogenèse et
la spermatogenèse.
1.2.1 L'oogenèse
1.2.1.1. Chez la femelle de nos animaux domestiques
L'oogenèse est le processus de la formation, de la croissance et de la maturation du
gamète femelle. Ce processus débute pendant la vie embryonnaire, continue après la
naissance avec une accélération au moment de la puberté et atteint son apogée au moment de
l'ovulation. Il ne peut être distingué de l'évolution du follicule ovarien au sein duquel il se
déroule en grande partie. L'évolution du follicule, à son tour, est indissociable de l'activité
cyclique de l'ovaire (ou cycle ovarien) à laquelle elle contribue par sa participation à un
contrôle endocrinien particulièrement perfectionné.
La phase de multiplication : elle est limitée dans le temps. Elle débute durant la vie
embryonnaire (phase de migration des CGP) et se termine avant ou peu après la naissance
(voir tableau 1.1). La transformation des oogonies en oocytes I suit immédiatement la phase
de multiplication et a également une durée déterminée (voir tableau 1.1). La première
division méiotique (DM1) se déroule jusqu'au stade diplotène auquel succède une longue
phase caractérisée par une modification du noyau (les tétrades se décondensent, s'entremêlent,
un ou plusieurs nucléoles apparaissent, une membrane nucléaire se reforme : c'est le stade
dictyé). Le noyau prend le nom de vésicule germinative. La première prophase ne se
poursuivra qu'après plusieurs mois ou plusieurs années, au cours d'un cycle ovarien, après la
puberté.
La phase d'accroissement : elle se déroule au sein du follicule. L'oocyte I, au stade
dictyé, s'entoure d'une assise de cellules folliculeuses qui dérivent du mésonéphros. Ces
cellules constituent bientôt une enveloppe épithéliale aplatie, ébauche de la granulosa. Elle
est séparée du stroma ovarien par une membrane basale : la membrane de Slavianski.
L'ensemble constitue le follicule primordial (figure 1.5).
Les cellules folliculeuses prolifèrent pour former d'abord un épithélium cubique
monostrastifié puis une structure pluristratifiée : la granulosa. L'assise cellulaire la plus
interne conserve sa morphologie épithéliale cubique et prend le nom de corona radiata.
Pendant ce temps, l'ovocyte I entre en accroissement. Dans l'interstice qui le sépare de sa
corona radiata se dépose une couche mucoprotéique formée conjointement par l'ovocyte et les
cellules de la corona radiata : la zone pellucide. A ce stade, le follicule est qualifié de
follicule primaire (voir figure 1.6).
Pendant son accroissement, l'ovocyte I conserve d'étroites relations avec les cellules de
la corona radiata (figure 1.6). Celles-ci envoient au travers de la zone pellucide de fins
prolongements cytoplasmiques dont les extrémités renflées établissent avec l'ovocyte des
relations jonctionnelles (desmosomes et gap junctions).

Ces jonctions permettraient aux cellules de la corona radiata de céder à l'ovocyte divers types
de matériaux nutritifs nécessaires aux premières divisions de l'embryon (accumulation de
ribosomes, d'ARNm, d'ARNt et des précurseurs nécessaires à la synthèse des
macromolécules).
Autour de la granulosa pluristratifiée, les cellules du stroma ovarien se condensent en
une couche préthécale. Au terme de l'évolution du follicule primaire, l'ovocyte I a presque
atteint son diamètre maximum. Les follicules primordiaux et primaires constituent le groupe
des follicules préantraux (ou précavitaires).
La phase de maturation: La prolifération des cellules de la granulosa se poursuit
activement, augmentant le diamètre du follicule. L'ovocyte I atteint son diamètre maximum
de 80 m (souris), 120 m (Femme), 150 m (vache). A son côté, au sein de la granulosa
apparaît une cavité ou antrum formée par la coalescence d'inclusions expulsées par les
cellules folliculeuses. Le follicule prend le nom de follicule secondaire ou antral. La
condensation des cellules du stroma ovarien autour du follicule se différencie en deux couches
distinctes: la thèque interne de nature épithélioïde entourant la membrane de Slavianski et la
thèque externe de nature conjonctivo-musculaire (figure 1.5). Des vaisseaux sanguins
colonisent les deux thèques, mais ne traversent pas la membrane de Slavianski, laissant la
granulosa momentanément avascularisée. Pendant ce temps, l'antrum se dilate par
accumulation d'un liquide (liquor folliculi) d'origine complexe (sécrétions de la granulosa, des
deux thèques et exsudation plasmatique). La granulosa tend à s'aplatir et l'ovocyte est refoulé
à un pôle de l'antrum, entouré de sa zone pellucide et de quelques assises de cellules
folliculeuses. L'ensemble, qui fait saillie dans l'antrum, prend le nom de cumulus oophorus
(figure 1.5).
L'étape finale de la croissance folliculaire est le follicule mûr ou préovulatoire
dénommé follicule de de Graaf. Son diamètre varie suivant l'espèce : - 15-20 mm pour
les grandes espèces (Femme, jument, vache).
- 5-8 mm pour les espèces moyennes (brebis, chèvre, truie).
- 2-3 mm chez les carnivores
- 0,1-1 mm chez la lapine et les rongeurs (rat, souris, hamster)
Les follicules occupent la corticale de l'ovaire, la médullaire étant formée d'un stroma
conjonctif richement vascularisé et de cordons épithéliaux (rete ovarii). Les follicules
primordiaux se localisent immédiatement sous l'enveloppe conjonctive de l'ovaire (ou
albuginée). A mesure qu'ils grandissent, ils s'enfoncent plus profondément dans le cortex. Du
fait de sa croissance, le follicule de de Graaf refait saillie à la surface de l'ovaire. C'est à cet
endroit qu'il se rompra pour libérer l'ovocyte au moment de l'ovulation (figure n° 1.5).
La croissance folliculaire est un processus continu. Elle débute durant la vie fœtale,
voire peu après la naissance (lapine) mais dépasse rarement le stade de follicule primaire. En
effet, tous les follicules qui ont entamé leur croissance avant la puberté sont voués à l'atrésie
(dégénérescence par apoptose) avant d'atteindre la stade antral. Il y a donc une perte très
importante de cellules germinales qui réduit considérablement le stock non renouvelable de
cellules germinales disponibles au début de la puberté pour assurer la reproduction de l'espèce
(figure n° 1.7).

A la puberté, sous l'influence des gonadotropines hypophysaires (Follicle Stimulating
Hormone ou FSH et Luteinizing Hormone ou LH) s'installe le cycle ovarien ou œstral, qui ne
s'interrompra que durant la gestation (ou à la ménopause chez la Femme). Ce cycle a une
durée variable suivant les espèces (voir tableau 1.3) et se décompose en deux phases,
folliculaire et lutéale, séparées par l'ovulation. En phase folliculaire, un grand nombre de
follicules s'accroissent - processus dont on a vu qu'il est continu - mais la plupart sont voués à
l'atrésie. Un seul d'entre eux chez les espèces monotoques (un foetus) ou quelques-uns chez
les espèces polytoques (plusieurs fœtus) iront jusqu'au terme de leur maturation et ovuleront.
L'ovulation (ou ponte ovulaire) est déclenchée par une décharge de LH. Durant les heures qui
précèdent l'ovulation, l'ovocyte I achève sa première division méiotique ou de maturation
(DM1). Au niveau cytoplasmique, se produit une redistribution des organites cellulaires ainsi
qu'une augmentation du nombre de granules corticaux qui migrent en périphérie.
Enfin les prolongements cytoplasmiques des cellules de la corona radiata se rétractent
progressivement et les contacts avec l'ovocyte sont rompus. L'ovocyte est désormais isolé au
sein de sa zone pellucide. Au niveau nucléaire, la vésicule germinative se rompt (Germinal
Vesicle BreakDown ou GVBD) et les tétrades migrent sur le premier fuseau achromatique.
La télophase I aboutit à une division très asymétrique de l'ovocyte I et se termine par la
séparation d'un ovocyte II de même taille que l'ovocyte I et l'expulsion d'un premier globule
polaire logé dans l'espace périvitellin, sous la zone pellucide (figure 1.8).
La seconde division méiotique (DM2) s'amorce immédiatement pour se bloquer, chez
la plupart des espèces, au stade de métaphase II. Elle ne s'achèvera qu'en cas de fécondation
(ou d'activation spontanée), aboutissant à la séparation d'un ootide de même taille que
l'ovocyte II et d'un second globule polaire. L'asymétrie de ces deux cytodiérèses a pour but de
produire une seule ootide à partir de chaque ovocyte I afin de ne pas disperser les réserves
cytoplasmiques accumulées durant la maturation. Notons que chez le chien et le renard, ces
deux divisions ont lieu après la fécondation (figure 1.8)
1.2.1.2. Chez la volaille
Les cellules germinales primordiales peuvent être histologiquement identifiées dans
le croissant germinal situé au niveau du pôle antérieur du blastodisque, en avant de la ligne
primitive, vers 18 heures d'incubation (stade 4 de Hamburger et Hamilton, voir figure 1.2).
Elles s'y multiplient par mitose et pénètrent activement dans les capillaires sanguins par
mouvements amoeboïdes (stades 4-8, 18-30 h). Arrivées au voisinage des ébauches
gonadiques, elles quittent activement les capillaires pour les coloniser (stades 20-24, 70-80
h). Chez la femelle, les PGC de la crête génitale droite migrent vers la crête génitale gauche
(stade 24, 95-100 h). Au sein de la crête génitale gauche, les oogonies continuent de se
multiplier.
Vers le 8ème jour d'incubation (stade 34) la crête génitale se différencie en ovaire et
les oogonies entrent en méiose pour se transformer en ovocytes I. Ces derniers seront
précisément arrivés au stade pachytène lors de l'éclosion du poussin. Ils évoluent ensuite vers
le stade diplotène (fin de prophase I) pour y rester bloqués pendant des mois ou des années
(stade dictyé). Durant cette période embryonnaire et post-éclosion, tous les ovocytes I
subiront un accroissement lent pour atteindre un diamètre de 100-200 m à l'éclosion et 1 mm
vers 4-5 mois, au moment de la puberté. La première division de maturation (DM1) reprendra
quelques heures avant l'ovulation, menant à la séparation de l'ovocyte II et du premier globule
polaire.
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%