SIDA ET DEFENCES IMMUNITAIRES Chapitre I : Le sida I) Le VIH

SIDA ET DEFENCES IMMUNITAIRES
Chapitre I : Le sida
I) Le VIH un rétrovirus
1) Un parasite intra cellulaire
Comment se présente le virus du VIH, ressemblance et différence avec une cellule ?
Doc 1 p 364 : les constituants essentiels du virus sont un acide nucléique l’ARN et des protéines, il ne possède ni
appareillage de synthèse de protéines, ni de systèmes producteur d’énergie.
Le virus ne peut donc se reproduire sans détourner le métabolisme d’une cellule.
Un virus est un agent susceptible d’infecter une cellule dans laquelle il se multiplie : c’est un parasite intra cellulaire.
En dehors de la cellule il apparait sous forme de particules virales de très petite taille. Ces particules comprennent une
coque protéique qui renferme le génome viral, le support est un acide nucléique. Ces particules assurent donc le
transport du génome d’une cellule à une autre.
2) Les rétrovirus
# doc 2 p 365
Le VIH apparient à un type de virus particulier : les rétrovirus, caractérisé par un matériel génétique de type ARN
associé à une enzyme essentielle à la multiplication intracellulaire du virus : la transcriptase inverse.la coque ou
capside protéique est pour le virus VIH entourée par une enveloppe formée d’une bicouche de lipides, dans laquelle
sont intégrés des protéines.
3) La pandémie et les modes de transmission.
# Doc 1et 2 p362, 363L’historique et extension de la pandémie avec une explosion de l’épidémie en Afrique sub-
saharienne.
Doc 3 : transmission par voie sexuelle de 72 % ou par voie sanguine ou de la mère à l’enfant.
Les particules virales sont transportées par les liquides sang, sperme et lait.
II) Les cellules cibles du VIH
1) L’infection par le VIH
# Doc 2 p365 : Il y a complémentarité de forme entre la protéine d’enveloppe virale GP 120 et le récepteur CD4
présent à la surface des cellules cibles, donc pour le virus la cellule cible possède des structures membranaires
reconnues par le virus et auquel il est capable de se fixer.
L’entrée dans le virus fait intervenir la protéine membranaire sur laquelle le virus s’amarre, on retrouve cette protéine
sur d’autres cellules immunitaires, les macrophages, les lymphocytes T4 et les monocytes.
Les macrophages peuvent constituer de véritable réservoir de virus , surtout dans les ganglions lymphatiques.
2) La multiplication des particules virales
Comment se multiplie les particules virales ? activité 3p366
# Doc 1 et 2 p 366,367 : Le virus se propage dans l’organisme par le sang et la lymphe
Pour se multiplier le virus détourne l’activité de la cellule hote. La transcriptase inverse et l’intégrase vont permettre la
production d’un provirus intégré dans l’ADN du LT4. Les ARN viraux produits par la transcription des provirus ne
pourront être traduits en protéines virales que par les ribosomes de la cellule hôte. C’est aussi cette dernière qui va
fournir l’ATP nécessaire à l’assemblage des acides aminés pour produire les polypeptides viraux.
3) Les conséquences immunologiques de l’infection.

# Doc 4 p363 : dans les semaines qui suivent l’infection le nombre de lymphocytes T4 chute brutalement, puis assez
rapidement leur valeur retrouve un taux proche de la normal. C’est le signe d’une réaction des défenses immunitaires.
En même temps on observe une augmentation de la concentration des anticorps dirigés contre les protéines de
l’enveloppe du virus ( antigènes) et une augmentation du nombre de lymphocytes T cytotoxiques, capables de lyser les
cellules infectées par le virus.
Le sida est caractérisé par trois phases la première : primo-infection peut ressembler à une maladie virale bénigne.
Puis une phase asymptomatique de durée variable suivant les individus et en fonction d’un traitement éventuel.
Jusqu’à 12 ans latence.
Le troisième stade : le sida confirmé est marqué par l’apparition de maladies opportunistes.
Pendant la phase asymptomatique le nombre de lymphocytes T4 baisse progressivement. Le virus est très peu présent
dans le sang. La propagation du virus se fait de cellule à cellule par la formation de syncytium.
Le VIH y reste inactif et donc invisible pour le système immunitaire mais en se multipliant dans les cellules il
constitue de véritables réservoirs.
Quand la concentration en lymphocyte T4 devient inférieure à 200 cellules par microlitre de plasma la maladie se
déclare.
La concentration d’anticorps contre les protéines virales diminue brutalement ainsi que le nombre de lymphocytes T
cytotoxiques.
La structure des ganglions s’altère, des maladies opportunistes se développent à cause de l’immunodéficience
croissante.
Doc 3 p 373
Le VIH s’est attaqué à deux population cellulaires essentielles au bon déroulement de la réponse immunitaire, les
lymphocytes T4 et les macrophages.
III) Quelques caractéristiques essentielles de la réponse immunitaire contre le VIH
1) La séropositivité
# TP Le test ELISA
# doc 1p368 :une réaction colorée ne peut apparaitre dans un puit de la plaque que si des anticorps anti
VIH sont présents dans le sérum à tester. Les anti-anticorps marqués par une enzyme E reconnaissent de manière
spécifique les anticorps anti –VIH et se fixent sur ces derniers.
Le malade produit des anticorps, la production de ces molécules définit la séropositivité du patient. Les anticorps se
fixent sur de très nombreuses protéines du virus VIH et empêche ainsi sa pénétration dans les lymphocytes et les
macrophages.
Mais les anticorps ne peuvent pas agir conter les cellules déjà infectées qui deviennent de véritables usines à produire
de nouvelles particules virales.
2) La production de lymphocytes cytotoxiques
Les lymphocytes T8 se transforment en lymphocytes cytotoxiques capables de reconnaitre les cellules infectées et de
les détruire.
Ces réponses sont dites spécifiques et impliques un certain délais entre la contamination et leur mise en place . C’est
pour cela qu’il existe un délai entre la séropositivité détectable et la primo infection qui peut aller de 1 à 3 mois.
# Doc p370, 371 : Le western Blot est plus précis que le test ELISA ou il peut y avoir de faux positifs.
Il s’agit ici aussi d’une réaction immunoenzymatique, les sujets11, 12 et 13, sont positifs au VIH1 car leur sérum
contient des anticorps dirigés contre les protéines d’enveloppe du virus GP 160 et GP 120 et contre au moins une
autre protéine du VIH1. Le sujet 10 est suspecté positif au VIH2, la seule bande P31 est insuffisante pour une
positivité au VIH1.

Chapitre 2 Les mécanismes de l’immunité
Rappel du collège : la peau et les muqueuses constituent une première ligne défense. Quand un antigène pénètre dans
l’organisme il entraine une première réponse immédiate, les microorganismes sont alors phagocytés par les
macrophages et les différents polynucléaires.
Puis une seconde réponse immunitaire se met en place.
Quelles sont les propriétés des anticorps ?
I) Les anticorps effecteurs de la reponse immunitaire acquise
# Doc 2et 3 p 388,389 : comment mettre en évidence la réaction antigène –anticorps, la présence d’arcs de
précipitation entre le puits centrale et les différents puits périphériques s’explique par la rencontre, suite à la migration
à l’intérieur du gel, d’un anticorps et de l’antigène correspondant. L’absence d’arcs au niveau des autres puits indique
que les substances qui se sont rencontrées à l’intérieur du gel ne se sont pas reconnues, et n’ont pas formées de
complexe immun.
Les complexes immuns forment en général des agrégats de relativement grande dimension, car la molécule
d’anticorps possède deux sites de fixation sur l’antigène, une molécule d’anticorps peut donc se fixer sur deux
antigènes différents et ainsi former des agrégats volumineux. Dans le cas de cellules on parle d’agglutination car en
présence d’anticorps spécifiques d’antigènes sur leur membrane, les cellules semblent se coller les unes aux autres
pour former un amas, les anticorps servant alors de liaison entre les différentes cellules.
# TP outcherlony et syphilis
1) La structure des anticorps
# TP anticorps et anagène
# Doc 1 une molécule d’ADN est constituée en moyenne de 1325 acides aminés.
- Les régions variables des deux chaines correspondent aux 100 premiers acides aminés environ
- Les régions constantes correspondent au reste de la séquence des deux chaines.
Etant donné les reploiements de la chaine polypeptidique, les régions hypervariables forment, à l’extrémité des bras Y
une région dont la forme tridimensionnelle peut varier d’un anticorps à l’autre en fonction de la séquence de ces
régions hypervariables : c’est le site anticorps. Pour un anticorps donné, ce site a une forme complémentaire de
l’antigène reconnu spécifiquement par cet anticorps.
Les anticorps plasmatiques sont des macromolécules de nature protéique. Elles appartiennent aux groupes des
immunoglobulines.
Les AA sont formés de 4 chaines 2 chaines lourdes et 2 chaines légères identiques 2à2 et liées par des ponts disulfures
.Chaque chaines comprends 2 parties, une partie constante et une partie variable.
Les parties variables sont spécifiques de chaque catégorie d’anticorps.
Les parties constantes des chaines lourdes aux extrémités terminales portent des sites liaisons non spécifiques avec
diverses cellules immunitaires, macrophages et lymphocytes B.
2) La liaison anticorps –antigène : le complexe immun.et son rôle.
# Doc 1 et 2 p 392, 393, ce système de phagocytose existe dès la naissance et est donc qualifié d’inné. Un phagocyte
est capable de reconnaitre un élément étranger et de le phagocyter puis de le lyser (digérer).
La liaison entre antigène et anticorps conduit à la formation de complexe immun. Ces complexes agglutinent
l’agresseur qui porte l’antigène .Ils accélèrent son élimination, en stimulant les phagocytes.
Cette propriété va être facilitée lorsque l’organisme aura développé des réactions immunitaires acquises aboutissant à
la synthèse d’anticorps, le phagocyte possédant sur sa membrane des récepteurs spécifiques de la partie constante des
immunoglobulines, la reconnaissance sera alors facilitée et donc plus rapide.
Les polynucléaires peuvent aussi se fixer sur les anticorps qui recouvrent un agresseur trop volumineux pour être
directement phagocyté, un ver parasite par exemple, et également secréter des enzymes qui détruiront l’agresseur.
Les anticorps agissent également contre les molécules solubles, les toxines, les bactéries exogènes ( ne pénétrant pas
dans les cellules) les parasites extracellulaires (vers) et les virus avant qu’ils ne pénètrent dans leurs cellules cibles.

II) L’origine des anticorps
1) Les plasmocytes
# Doc 1 et 2 p 394, 395 : en comparant la structure du plasmocyte et du lymphocyte B on observe un développement
de l’appareillage de synthèse des protéines dans le cytoplasme : réticulum endoplasmique granuleux, mitochondries et
appareil de golgi.
Un antigène déterminé qui pénètre dans l’organisme est détecté par un seul clone de lymphocyte B car les sites
anticorps d’une molécule d’immunoglobulines reconnaissent de manière spécifique un antigène donné.
Les immunoglobulines situées à la surface d’un lymphocyte B sont toutes identiques.
Il en est de même pour toutes les cellules d’un clone de LB puisque deux cellules issues par mitose d’une cellule mère
possèdent la totalité de l’information génétique de cette dernière.
On peut parler de sélection clonale puisque, en présence d’un antigène donné, un seul clone de LB va être activé.
.
Les anticorps du plasma sont produits par les lymphocytes B sécréteurs ou plasmocytes, ce sont des cellules de
grandes tailles 20 µm avec un cytoplasme abondant contenant beaucoup de réticulum endoplasmique, des
mitochondries et un appareil de golgi développé. Ils produisent et secrètent une grande quantité d’anticorps circulants.
Ils ne se divisent pas leur durée de vie est courte Ils produisent une seul variété d’anticorps.
Les immunoglobulines libérées par un plasmocytes possèdent exactement la même spécificité antigénique que les
anticorps membranaires du LB qui a reconnu l’antigène .Car c’est toujours le même gène d’immunoglobuline qui
s’exprime dans le plasmocyte. En fait la seule différence entre l’anticorps soluble et l’anticorps membranaire est que
ce dernier possède , à l’extrémité du Y , une séquence d’acides aminés hydrophobes qui lui permettront un ancrage
dans la membrane , séquence qui a disparu chez les anticorps solubles.
Avant tout contact avec des antigènes, l’organisme possède 10 8 lymphocyte B différents.
Les plasmocytes ne sont pas présents chez des personnes saines
2) La sélection clonale
L’introduction d’un antigène dans l’organisme conduit à la sélection clonale de quelques lymphocytes B, parmi
plusieurs dizaines de million.
Cela déclenche après quelques jours la sélection d’anticorps spécifiques : c’est la sélection clonale. Qui sera suivit par
la phase d’amplification et accompagné d’une mise en circulation de lymphocytes B mémoire.
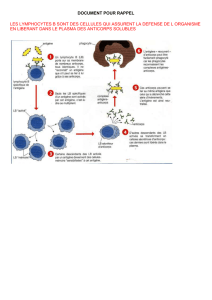
# Schéma bilan.
III) Les lymphocytes T , origine et destins
1) L’origine
# Doc p 398, 399, Les LB détectent les antigènes dans les liquides circulants alors que les LT les détectent sur les
membranes cellulaires. Dans les deux cas la détection se fait grâce à des immunoglobulines implantées dans la
membrane du lymphocyte
- Dans le cas du LB , il s’agit d’anticorps membranaires formés de quatre chaines polypeptidiques et présentant
deux sites de reconnaissances de l’antigène.
- Dans le cas du LT il s’agit d’un récepteur T formé seulement de deux chaines polypeptidiques et présentant un
seul site de reconnaissance.
Les lymphocytes T8 se forment dans la moelle osseuse .Ils acquièrent leurs récepteurs spécifiques T dans le thymus
puis ils gagnent les ganglions lymphatiques
2) Le mode d’action
# Doc 3 p 399, Un LT8 activé par un antigène, engendre un clone de LT8, qui vont tous se différencier en LTc,
contrairement à ce qui se passe pour les LB il n’y a pas ici de LTc mémoire.
# Doc 396,397 l’activité cytotoxique se fait en trois temps 1) liaison à la cellule cible2) une phase dépendant du Ca2+
qui modifie le contenu de la cellule cible 3) phénomène d’apoptose (mort de la cellule).

Certains Lt cytotoxiques contiennent de la perforine, une protéine capable de former des pores dans les membranes.
En présence de calcium les monomères de perforine se lient à la membrane de la cellule cible et se polymérisent pour
former des canaux transmembranaires. La cellule cytotoxique survit et peut continuer à lyser d’autres cellules.
D’autres phénomènes de lyse existent, des cytokinines, se fixent à des récepteurs membranaires et les cellules cibles
sensibles aux cytokinines meurent.
Les cellules infectées par un virus possèdent à leur surface des fragments peptidiques issus des protéines de l’agent
infectieux.
Les lymphocytes T cytotoxiques possèdent des récepteurs T présents à leur surface et ils vont se lier avec un fragment
peptidiques .La reconnaissance déclenche la sécrétion, exocytose de protéines, les perforines, qui vont former un
canal qui va permettre des échanges d’ions et qui va conduire à la lyse de la cellule.
IV) Les lymphocytes T4, pivots des réactions immunité acquise
1) L’activation des LT4
# Doc p400,401 l’irradiation détruit toutes les cellule souches des cellules immunitaires dans la moelle osseuse. La
privation de thymus empêche la production de lymphocytes T.Il y a nécessité d’une coopération cellulaire entre
lymphocytes B et T pour que cette production soit efficace.
La coopération se fait par un agent soluble.
La pénétration d’un antigène dans l’organisme entraine l’activation de LT4 spécifique de cet antigène, cette activation
est liée à la présentation de l’antigène par un macrophage.
Les LT4 spécifique par leurs récepteurs T se fixent, ils vont former un clone de cellule. Ils sont sélectionnés et
deviennent sécréteurs de messagers chimiques les interleukines.
2) La stimulation de LB et de LTc
Des ganglions, les clones de LT4 activés produisent des interleukines qui activent les LT8 et le LB possédant des
récepteurs spécifiques de l’antigène et des récepteurs aux interleukinesIL2.
Les LT8 et les LB se multiplient et se différencient en LTc ou en plasmocytes sécréteurs d’anticorps
Les LT4 jouent un rôle central. Des LT4 mémoire se forment
SCHEMA BILAN
Les vaccins et la mémoire immunitaire
I)La mémoire immunitaire
1) Les vaccins
1
/
5
100%