DESPREZ Christophe KHAN Pierre 27/01/11 Périconceptologie

1/13
DESPREZ Christophe
KHAN Pierre
27/01/11
Périconceptologie, Axe HHO, Dr K.Morcel
AXE HYPOTHALAMO-HYPOPHYSO-
OVARIEN
Introduction
Il existe des fluctuations hormonales au niveau de l'axe hypothalamo-hypophysaire-ovarien (=
HHO) qui ont des actions sur :
l'ovaire ce qui va permettre la phase terminale de la folliculogénèse
l'ovulation par la décharge ovulante
sur la préparation de l'utérus à l'implantation de l'embryon en faisant croitre et maturer
l'endomètre par l'œstradiol et la progestérone
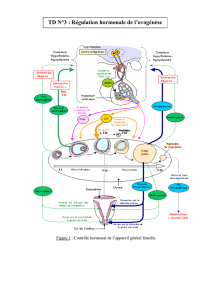
Il y a des interactions complexes entre le système hypothalamo-hypophysaire et l'ovaire. Il y a 3
types de régulation :
la régulation neuro-endocrine, du complexe HH jusqu'à l'ovaire et vice versa
la régulation paracrine au sein même du tissu ovarien, entre les cellules de l'ovaire
la régulation autocrine dans les cellules elles-mêmes
L'objectif de ces interactions est d'obtenir une croissance folliculaire qui permettra l'ovulation, la
fécondation et la grossesse.
I – L'hypothalamus
C'est une glande, une partie du diencéphale qui forme le plancher et les parois du 3ème ventricule à la
base du cerveau. Il reçoit des informations nerveuses et chimiques. C'est l'hypothalamus antérieur
qui nous intéresse ici, il va moduler l'hypophyse.
Il contient des noyaux formés de cellules neuro-endocriniennes qui ont des propriétés de neuro-
sécrétion. Ces noyaux sont sensibles :
• aux neuro-médiateurs venus par voie nerveuse
• aux hormones véhiculées par voie sanguine sécrétées par les ovaires
Ils élaborent des substances de type neuro-hormones qui migrent dans des petits grains le long de leur
axone jusqu'à sa terminaison sur une branche du système porte ou système de Poppa.
Ce système est un réseau vasculaire joignant l'hypothalamus à l'hypophyse antérieur. ( voir schéma à
la fin )

2/13
L'hypothalamus a une action sur l'hypophyse par l'intermédiaire d'une neuro-hormone :
= LH-RH (LH Releasing Hormone)
= GnRH (Gonadotropin Releasing Hormone)
= Gonadoréline
cette neuro-hormone va stimuler l'hypophyse qui va libérer les gonadotrophines (LH et FSH).
La GnRH (ou LH-RH) est sécrétée en permanence et de façon pulsatile; pourtant elle n'est pas
retrouvée dans un prélèvement sanguin. En effet elle reste dans le système porte ( de Poppa ) et ne
se diffuse pas dans la circulation générale.
A – La GnRH
Cette neuro-hormone est un décapeptide.
1 – Les Neurones à GnRH
Les neurones à activité sécrétoire GnRH sont localisés essentiellement dans l'hypothalamus au
niveau du noyau arqué. Les neurones à GnRH sont originaires de la placode olfactive et migrent
secondairement dans le cerveau au cours de l'embryogenèse.
NB : Il y a donc des pathologies qui associent des anomalies de l'olfaction et des anomalies du cycle
menstruel comme le syndrome de Kallmann de Morsier = dysplasie olfactogénitale qui conjugue
anosmie et trouble hormonale du cycle menstruel.
2 – La Sécrétion
La GnRH a une demi-vie courte de 2 à 4 minutes.
La GnRH doit avoir une sécrétion pulsatile. Ces pulsations doivent avoir une fréquence et une
amplitude variable et ceci est modulé par les stéroïdes gonadiques par un rétro-contrôle des
hormones ovariennes (œstrogène et progestérone).
En cas sécrétion continue de GnRH, il y a désensibilisation des récepteurs à GnRH, l'hypophyse est
ainsi bloqué et les sécrétions de LH, FSH s'estompent.
La variation de fréquence module la réponse hypophysaire et le rapport FSH/LH. Il y a des
pathologies où le rapport est inversé.
➝ Quand la fréquence de libération de GnRH augmente, le rapport FSH/LH diminue
➝ donc le LH augmente et la FSH diminue (et vice-versa).
Il y a des moments du cycle où c'est la LH qui va avoir besoin d'être sécrétée. La LH c'est ce qui
permet l'ovulation. Donc en pré-ovulatoire il va y avoir une sécrétion de LH, ce qui nécessite une
augmentation de la fréquence des pulses de GnRH. Certaines patientes n'ovulent pas car elles n'ont
pas une fréquence assez importante de GnRH.
Phase du cycle
Période (min) entre chaque pulse de GnRH
Folliculaire:
Début :toutes les 94 min
Fin : 76
Lutéale:
Début : 103

3/13
Fin : 216
3 – Les Récepteurs à GnRH
Ces récepteurs sont situés de l'anté-hypophyse.
En fin de phase folliculaire, il y a besoin de moins de FSH.
Il faut des récepteurs à la GnRH qui sont des protéines membranaires. Ces récepteurs sont calcium-
dépendants. Ils permettent deux types d'action après fixation de GnRH :
à court terme: entrée de calcium dans la cellule qui va se lier à la calmoduline ce qui va
engendrer l'exocytose des grains de sécrétion de FSH et LH
à long terme: ils agissent sur la transcription des gènes des sous-unités des hormones
gonadotrophines par activation de la protéine kinase C.
Le caractère pulsatile est nécessaire à une réponse hypophysaire normale. L'administration
continue de LH-RH bloque les canaux calciques et arrête le fonctionnement des récepteurs ce qui
diminue leur nombre. En effet dans la régulation autocrine, chaque hormone régule la formation de
son propre récepteur.
Il y a plusieurs contrôles. De nombreuses afférences intègrent les informations venues de tout le
système nerveux (puisque c'est un système neuro-endocrinien) et interviennent sur la terminaison
portale des neurones neuro-sécréteurs. Il y a des influences stéroïdo-sensibles : l'oestrogène et la
progestérone sécrétées par l'ovaire vont faire un rétro-contrôle sur les neurones de l'hypothalamus.
B – Régulation de l'hypothalamus
1- Phénomène de feed-back :
Les taux circulants de stéroïdes sexuels (œstrogènes, androgènes ou progestatifs), quand ils
atteignent un certain seuil, vont freiner la sécrétion de GnRH = feed back négatif. Inversement,
quand ils sont bas, ils vont la stimuler = feed back positif.
Le taux de LH est réglé par les stéroïdes circulants. La progestérone diminue la fréquence des
pulsations de GnRH et augmente leur amplitude, ce qui va diminuer la LH plasmatique. La
progestérone est présente pendant la phase lutéale, l'ovulation est faîte donc il n'y a plus besoin de
LH. L'œstradiol, qui augmente progressivement pendant la phase folliculaire, va activer la sécrétion
de GnRH et modifier la sensibilité hypophysaire à la GnRH.
Le taux de FSH est modulé par l'inhibine A sécrétée par les cellules de la granulosa qui agit au
niveau des cellules hypophysaires gonadotropes sur des récepteurs spécifiques autres que les
récepteurs à GnRH.
2 - Action du SNC :
Il y a une connexion entre l'hypothalamus et le SNC. Le rôle de la dopamine, des prostaglandines et
des endorphines est en cours de recherche.

4/13
La régulation de la sécrétion de LH-RH est assurée par :
des interactions :
longues entre les ovaires et l'hypothalamus (par l'intermédiaire des stéroïdes).
courtes avec l'hypophyse et ses gonadotrophines.
ultra-courtes au sein même de l'hypothalamus (le LH-RH est capable d'inhiber sa
propre synthèse).
des modulations :
inhibition par le système dopaminergique : neurone à dopamine dans le noyau
arqué qui inhibe la production de GnRH, notamment chez les jeunes filles
sportives souvent en aménorrhée car on a une diminution de la GnRH .
stimulation par le système noradrénergique : neurone à NA et sérotonine dans le
mésencéphale.
Actions variables d'autres facteurs : peptides cérébraux, facteurs de croissance,
endorphines...
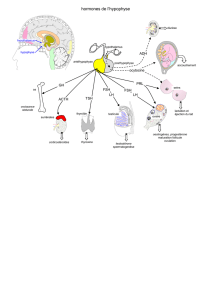
II – L'hypophyse
C'est la 2ème glande du système. Au niveau de l'hypophyse antérieure, les mêmes cellules
sécrètent les gonadotrophines ou gonadostimulines : LH et FSH = hormones à polarité gonadique.
Leur sécrétion suit le mode pulsatile de la GnRH.
Il y a d'autres hormones sécrétées par l'anté-hypophyse : la TSH, les hormones de croissance, GH et
l'ACTH pour les hormones corticotropes. Elles vont chacune stimuler des axes différents.
A – Les gonadotrophines : LH et FSH
Ce sont des dimères glycoprotéiques, elles sont constituées de deux sous-unités différentes (α
et β) chacune formée d'une séquence de polypeptide porteuse d'une ou plusieurs chaînes
polysaccharidiques reliée de manière non covalente.
La sous-unité α est commune à toutes les hormones anté-hypophysaires.
La sous-unité β est spécifique à chaque hormone.
La LH a une demi-vie courte de 6h. Elle a une action très rapide.
La FSH a une demi-vie longue de 36h.
Les deux hormones sont sécrétées par les mêmes cellules de l'anté-hypophyse : les cellules
gonadotropes.
Les deux hormones agissent en synergie au cours du cycle menstruel. Elles sont stimulées par les
sécrétions pulsatiles de GnRH. Leur sécrétion est pulsatile parallèlement à celle de la LH-RH.

5/13
B – Régulation des gonadotrophines
1 – Interactions
longues avec l'ovaire par l'intermédiaire des stéroïdes et de l'inhibine qui ont une action
de feed back négatif
courtes avec l'hypothalamus, action variable de la GnRH en fonction des pulses.
ultra-courtes au sein de l'anté-hypophyse avec les autres cellules de l'anté-
hypophyse : lactotropes, thyréotropes et corticotropes respectivement pour la prolactine, la
TSH et l'ACTH. Action inhibitrice de ces cellules.
2 - Action de la FSH (Follicule Stimulating
Hormone ou hormone folliculo-stimulante)
Elle agit par l'intermédiaire de récepteurs spécifiques à la FSH situés sur les cellules de la granulosa
qui sont les seules à porter ce type de récepteurs au niveau du follicule ovarien. Elle est la seule a
avoir un rôle dans le développement et le recrutement du follicule.
Elle a une double action :
prolifération : elle permet la multiplication des cellules de la granulosa qui permet la
croissance du follicule.
On appelle ça une action dose-dépendante. La sensibilité des follicules à la FSH dépend
du nombre de récepteurs qui est proportionnel au nombre de cellules de la granulosa.
Il y a une notion de seuil de réceptivité qui va permettre les mécanismes de recrutement
et de sélection des follicules dont le seuil de sensibilité est suffisant. Seul le follicule le
plus sensible (donc le plus gros) sera apte à poursuivre sa croissance malgré la baisse de
FSH en fin de phase folliculaire.
différentiation : elle permet d'acquérir les fonctions des cellules de la granulosa :
- stéroïdogénèse (sécrétion des œstrogènes),
- synthèse d'inhibine,
- expression des récepteurs à la LH, il faut que la cellule soit sensible à FSH pour avoir
des récepteurs à la LH.
3 – Action de la LH (Luteinizing Hormone ou
hormone lutéo - stimulante)
Elle agit par l'intermédiaire de récepteurs spécifiques à la LH situés au niveau de la thèque interne,
de la granulosa puis du corps jaune. Les récepteurs de la granulosa apparaissent sous l'action de la
FSH vers le 10ème jour du cycle (follicule > 10mm).
Elle a plusieurs rôles :
le contrôle de la sécrétion des stéroïdes ovariens :
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%