6° partie : Immunologie (4 semaines)

Immunologie
Rappels 3ème (livre p 360-361):
- microorganismes susceptibles d’être infectieux
- système immunitaire :
Le SIDA (Syndrome d’ImmunoDéficience Acquise) est une maladie causée par un virus (VIH : Virus de
l’Immunodéficience Humaine) que le système immunitaire n’arrive pas à combattre. Comment peut-on expliquer
les différentes phases lors d’une infection par le VIH ? Comment peut-on expliquer l’évolution de la charge
virale au cours de la maladie ? Quelles connaissances du système immunitaire peut-on tirées de l’étude de ce
virus ?
Evolution de la charge virale de la contamination à la mort de l’individu
Chapitre 1 : Augmentation de la charge virale et primo-infection
Le SIDA est une maladie qui a été clairement identifiée en 1986 par le Professeur Montagnier (Institut Pasteur).
Les premiers cas repérés remontent à 1981. En 2004, on estimait à 44, 3 millions le nombre de personnes
séropositives (qui possèdent des anticorps anti-VIH dans leur sang), dont 65 % vivent en Afrique. Cette épidémie
planétaire est qualifiée de pandémie. 1 à 2% de la population mondiale est actuellement touchée.
La première phase de la maladie qualifiée de primo-infection est caractérisée par un pic de la charge virale
(quantité de virus mesurable dans le sang). Cette première phase dure 6 à 8 semaines.
Comment peut-on expliquer l’augmentation de la charge virale ?
Hypothèse : le virus se multiplie dans l’organisme.
I. Le V.I.H. : virus de l’immunodéficience humaine (http://www.snv.jussieu.fr/vie/dossiers/SIDA/index.htm)
A. Modes de transmission
Le virus du SIDA peut être transmis de diverses manières, qui impliquent différents fluides corporels : le sang, les
sécrétions génitales, le lait.
- Transmission par voie sexuelle (70 à 80 % des cas d'infection)
Le virus est présent dans les sécrétions génitales, et peut donc être transmis lors d'un rapport sexuel, qu'il soit
homosexuel ou hétérosexuel. Certaines maladies sexuellement transmissibles, et surtout la multiplication des
partenaires (sans protection lors des rapports) favorisent cette transmission.
- Transmission par le sang
Le virus étant présent dans le sang, il peut être transmis lors de tout "don" de sang d'un individu à un autre : lors de
pratiques toxicomanes (échanges de seringues), de manière accidentelle, ou lors de transfusions. Un dépistage
systématique des dons du sang a permis de réduire ce dernier mode de transmission (risque résiduel estimé à
1/500.000).
- Transmission materno-foetale
Le virus est capable de traverser la barrière hémato-placentaire, et ainsi de contaminer, in utero, un foetus. Le cas le
plus fréquent semble être toutefois lors de l'accouchement. De plus, le virus se retrouve dans le lait maternel, d'où
une contamination lors de l'allaitement (cas fréquent surtout en Afrique). Sans traitement, le VIH-1 se transmet à
VIH-1

15-20% de la mère à son enfant (30% si allaitement). Avec traitement préventif, le taux de transmission du VIH-1
baisse à moins de 8% (moins de 2% en Europe). Chaque jour, environ 1000 enfants naissent en Afrique porteurs du
VIH... La moitié de ces enfants décèdera des suites de la maladie dans les 2 premières années de vie.
B ; Carte d’identité du VIH
1) Généralités
Un virus est un parasite intracellulaire obligatoire ne pouvant se multiplier qu'à l'intérieur d'une cellule hôte et
utilisant sa machinerie cellulaire. Il contient : une information génétique (sous forme d'ADN ou d'ARN), et une
structure de protection souvent protéique, compacte, pour protéger son Acide Nucléique (La Capside). Il existe de
nombreux types différents de virus, parmi lesquels on retrouve en particulier les rétrovirus. Les rétrovirus sont des
virus d’un diamètre de 110 à 125 nanomètres, très répandus dans le monde animal. Ils sont la cause de différentes
formes de cancers, d’immunodéficiences, dont le sida, et de dégénérescences du système nerveux central. Leur
génome s’intègre sous forme d’ADN dans celui de la cellule hôte, pour ensuite s’exprimer pendant toute la vie
active de la cellule. Les lentivirus (ou Lentivirinae) font partie de cette famille : ces virus sont responsables de
pathologies à évolution lente.
L'exemple le plus connu de tels virus est le virus du SIDA : le VIH. Il s'agit d'un virus possédant un génome sous
forme d'ARN, contenu dans une capside protéique, elle même entourée par une membrane lipidique. On distingue
actuellement deux types de VIH : le VIH-1 et le VIH-2. Ces deux virus sont très proches (42 % d'homologie au
niveau de leur génome). Le VIH-1 est le plus répandu : c’est celui que l’on étudiera.
2) Structure du VIH (voir schéma)
gp 120 : protéine mbr portée par la bicouche lipidique, elle intervient dans la fixation du VIH aux LT CD4+ : elle
est capable de se fixer aux R CD4 de ces cellules (ainsi qu’à leur co-R). Issue de gp160 plus grande clivée par une
protéase cellulaire (de la cellule infectée).
gp 41 pr transmbr, elle intervient dans la fusion des mb du VIH et du LT4 infecté. Elle s’insère dans la mb des
lymphocytes après fixation de gp120 au récepteur CD4. Elle est issue d’une pr + grande (gp 160) clivée par une
protéase cellulaire.
Mb plasmique : bicouche lipidique taillée dans celle du lymphocyte lors du bourgeonnement.
Capside externe : répétition de p17gag (sous unité pr), protection du matériel génétique viral (issue d’un
précurseur gagp55 clivé par une protéase virale).
Capside interne : répétition de p24gag (sous unité pr), protection du matériel génétique (issue d’un précurseur
gagp55 clivé par une protéase virale).
Ce virus a la particularité d’avoir deux capsides protéiques.
La membrane plus les capsides protéiques forment l’enveloppe virale.
p7 gag et p9 gag : pr associées à l’ARN viral
Réverse Transcriptase ou Transcriptase inverse: ADN polymérase permettant la synthèse d’une molécule ADN
double brin à partir de l’ARN viral dans la cellule hôte (synthèse d’un brin d’ADN complémentaire à l’ARN viral
puis synthèse du deuxième brin d’ADN), le phénomène est appelé rétrotranscription.
Matériel génétique : ARN viral simple brin présent en double exemplaire.
Le génome du virus du SIDA se compose d'un ARN simple brin de 9181 nucléotides. Il comporte trois gènes
principaux (Gag, Pol, et Env), ainsi que quelques gènes de régulation, de petite taille. Il comporte de plus des
séquences spécifiques, situées à ses extrémités (5'UTR et 3'UTR - UTR = région non transcrite "UnTranscribed
Region"). Une fois rétrotranscrit sous la forme d'un ADN double brin (voir cycle), il s'exprime par le biais de deux
ARN messagers, qui aboutissent à la synthèse de trois protéines. Ces protéines sont ensuite clivées par des
protéases, pour aboutir aux différentes protéines virales (voir doc)
Le VIH (virus de l’immunodéficience humaine) appartient à la catégorie des rétrovirus (virus à ARN).
Comment l’ARN peut-il permettre la multiplication du virus ?
II. LES DIVERSES CELLULES PARASITEES PAR LE V.I.H.
A. Les cellules cibles
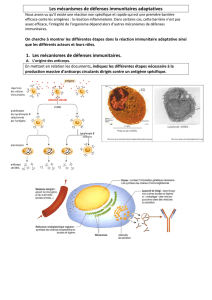
Les cellules cibles du virus sont des cellules du système immunitaire, c’est-à-dire de la famille des « globules
blancs » : les leucocytes. Comme les autres cellules sanguines, ils sont produits par la moëlle osseuse.
Il existe différents types de leucocytes :
- les macrophages (quand ils sont immatures, c’est-à-dire dans le sang, ils sont appelés monocytes), possèdent
trois fonctions principales : une activité de sécrétion (production de cytokines : molécules signal du système
immunitaire, régulant la production des globules, l’hémostase...) ; une coopération cellulaire (c'est une cellule
présentatrice d'antigènes, relation avec les lymphocytes) ; et la phagocytose (ingestion de bactéries, levures, débris
cellulaires...).
- les lymphocytes : Variété de globules blancs du sang. Ils interviennent dans la réponse immunitaire. Ils sont de
deux sortes : les lymphocytes B (production d'anticorps, Bone) et les lymphocytes T (T4 médiation et T8
cytotoxiques,Thymus, immunité à médiation cellulaire).

Populations cellulaires infectées
par le VIH
Caractéristiques moléculaires des cellules cibles
Lymphocytes T4
Présence de l'antigène membranaire CD4 impliqué l'interaction avec une
protéine de l'enveloppe virale
Présence d'un corécepteur membranaire CXCR4 nécessaire pour l'entrée du
VIH dans les cellules et pour une infection efficace
Monocytes et macrophages
dérivés des monocytes
Présence de l'antigène membranaire CD4 impliqué l'interaction avec une
protéine de l'enveloppe virale
Présence d'un corécepteur membranaire CCR5 nécessaire pour l'entrée du
VIH dans les cellules et pour une infection efficace
Le VIH infecte également des cellules du système nerveux central : on a pu détecter des séquences d'ADN et
d'ARN virales dans le cerveau des enfants et des adultes atteints de SIDA, ce qui suggère que la réplication virale
s'y effectue.
Les cellules cibles du VIH sont principalement des cellules immunitaires : lymphocytes T4, monocytes et
macrophages.
Pourquoi le VIH pénètre-t-il dans ces cellules et pas dans les autres ?
La protéine virale gp120 portée par la membrane du VIH est capable de se lier à la protéine membranaire CD4 des
lymphocytes T4 et des monocytes/macrophages. Une autre protéine lymphocytaire intervient dans cette liaison : on
parle d’un co-récepteur (sa nature varie selon le type de cellule immunitaire : protéine CXCR4 pour LT4 et CCR5
pour monocytes/macrophages).
Les membranes des cellules et des virus sont constituées de lipides et d’éléments membranaires ou
transmembranaires (tels que les protéines CD4) : ce ne sont pas des structures figées. Les protéines « circulent » au
sein des lipides. C’est de cette manière que les protéines virales gp 41 peut se mouvoir jusqu’au lieu de liaison
entre le virus et la cellule immunitaire et passer dans la membrane lymphocytaire. gp 41 permet une fusion des
deux membranes : le virus entre dans la cellule (sans sa membrane).
Les deux capsides se dissocient et l’ARN viral en double exemplaire, la reverse transcriptase ainsi que les protéines
associées à l’ARN sont libérées dans le cytoplasme du lymphocyte.
La réverse transcriptase synthétise d’abord un ADNc (complémentaire de l’ARN viral) puis le brin d’ADN
complémentaire de l’ADNc pour obtenir de l’ADN double brin. Ce double brin d’ADN entre alors par les pores de
l’enveloppe nucléaire dans le noyau, se circularise et s’intègre à l’ADN lymphocytaire.
Dès lors, la machinerie cellulaire transcrit les informations génétiques apportées par le virus : des ARNm sont
synthétisés grâce à l’ARN polymérase lymphocytaire (le génôme du VIH possède une structure particulière qui
permet la fabrication de différents ARNm à partir d’une seule séquence nucléique). Les ARNm quittent le noyau
pour être traduits dans le cytoplasme selon les modalités vues en 1°S. Des processus de maturation complexes
conduisent à la traduction de différents précurseurs protéiques. Des protéases cellulaires et virales les clivent pour
former les différentes protéines participant à la structure du VIH.
Puis les différents composants du virus s’auto-assemblent : ARN, RT, et protéines.
Le virus possède sa double capside mais pas encore de membrane : il sort de la cellule par le même système de
liaison protéique qui lui a permis d’entrer.
Ce processus ne tue pas la cellule infectée qui continue à produire de nouveaux virus. Le virus libéré dans le milieu
extracellulaire circule le temps de rencontrer une autre cellule portant la protéine membranaire CD4. Et le cycle
recommence.
Les LT4, monocytes et macrophages possèdent des protéines membranaires auxquelles le virus s’amarre par
l’intermédiaire d’une protéine de son enveloppe (la plus importante. de ces protéines membranaires des
cellules immunitaires étant CD4), ce qui lui permet de pénétrer dans la cellule hôte.
B. Arrivée des virus aux cellules cibles.
Le VIH est un virus très fragile. En effet le virus survit très difficilement s'il se trouve en dehors de l'organisme
humain. => Impossibilité d'infection par le simple touché, par l'utilisation des W.C. publics, par la sueur, par la
nourriture, les assiettes, les verres, les couverts. L'infection par le VIH ne peut pas se produire lors d'activités et de
contacts quotidiens ordinaires tels que les poignées de main, les embrassades, les baisers, la tous, les éternuements,
les piscines publiques, les sièges de toilette, les draps de lit, la vaisselle, les aliments, les maringouins ou les
animaux. Donner du sang ne comporte aucun risque. Vous ne pouvez pas contracter le VIH en donnant du sang
puisqu'une nouvelle aiguille est utilisée pour chaque donneur.

Le VIH se transmet exclusivement par les sécrétions sexuelles et par le sang. Il est libérée par la cellule réservoir
(lymphocyte T4 ou monocyte/macrophage) dans le milieu extracellulaire. Etant donné que les cellules immunitaires
sont réparties dans tout l’organisme pour assurer la défense contre des agents pathogènes, le VIH est présent dans
tous les compartiments liquidiens de l’organisme : le sang, la lymphe (les organes lymphoïdes étant des réservoirs
de cellules immunitaires présentent de forte charge virale), les sécrétions sexuelles (sécrétions vaginales, liquide
séminal, sperme).
Rmq : les conditions régnant dans le tube digestif (milieu extérieur) ne permettent pas au VIH de survivre dans les
sécrétions (salive…), idem pour les larmes
(Déjà vu lors TP1) TRANSMISSION PAR VOIE SEXUELLE
Toutes les relations sexuelles, qu'elles soient vaginales ou anales, buccales, homosexuelles ou hétérosexuelles,
peuvent transmettre les VIH. La transmission des VIH se font, dans les couples hétérosexuels, aussi bien dans le
sens homme-femme que femme-homme. En raison de la plus grande fragilité des muqueuses, les relations anales
sont plus "infectantes" que les relations vaginales. Toutes les infections génitales (MST généralement) de l'un des
partenaires augmentent considérablement le risque de transmission de la maladie, quelles soient situées sur le
gland ou le vagin. Les contacts oraux-génitaux et oraux-anaux comportent eux-aussi des risques. De ce fait, il est
recommandé d'utilisé un préservatif (il en existe de différents goûts) lors d'une fellation, et d'utiliser un carré de
Latex, ou du papier alimentaire pour les cunnilingus, et anulingus.
TRANSMISSION PAR VOIE SANGUINE
La Transmission du VIH par le sang peut avoir lieu à différentes occasions :
Lors d’une transfusion ou de l'injection de produits sanguins. Mais celle-ci est devenue très rare depuis la nouvelle
règlementation qui est en application depuis le 1er Aout 1985.
Par des seringues et des aiguilles souillées. C'est en fait le problème des drogués qui utilisent du matériel déjà
servi. En revanche, dans le milieu médical, les seringues, les aiguilles ou autres instruments ne présentent plus
aucun risque, tant sont rigoureuses les précautions qui entourent leur usage.
Au cours de la grossesse. En effet le VIH peut se transmettre de la maman à son bébé, soit à travers le placenta
Le VIH est transmis par voie sexuelle, par voie sanguine ou au cours de la grossesse de la mère à l’enfant.
(On ne sait pas si le virus se transmet pendant la grossesse : on ne peut faire de ponction de sang foetal, on
risquerait de transmettre le virus à un fœtus séronégatif. La moitié des enfants qui naissent séropositifs
développent la maladie et décèdent dans les 2 premières années de leur vie.)
Comment débute la maladie ?
III. LE V.I.H. ET LA CELLULE CIBLE.
Une fois le cycle du virus initié, il peut rester en latence (on parle de provirus) ou bien sa multiplication est activée
(début de la transcription de l’ADN viral incorporé dans le génome de la cellule hôte). Des virus sont libérés dans
les liquides biologiques et identifiés comme corps étranger par le système immunitaire.
Lors de la primo-infection, le système immunitaire est assez efficace contre le virus. Des morceaux de VIH ou des
VIH entiers sont présents dans les différents liquides biologiques. Ils sont reconnus comme des antigènes par les
cellules du système immunitaire (Molécule étrangère à l'organisme, qui déclenche une réaction de défense
(réponse immunitaire) de ce même organisme, caractérisée par la production d'anticorps). Les lymphocytes B
reconnaissant spécifiquement des antigènes du VIH (ex ; morceaux de protéines virales) sont alors activés : ils se
multiplient de façon clonale et se mettent à produire des anticorps (Molécules de défense de l'organisme, protéines
(immunoglobulines) fabriquées par les lymphocytes (globules blancs du sang), en réponse à la présence d'un corps
étranger, l'antigène.) Les anticorps spécifiques sont libérés dans les liquides biologiques, ils se lient aux particules
virales. Cet entourage d’anticorps les conduit à se faire phagocyter par les macrophages la charge virale
sanguine diminue.
La fabrication d’anticorps prend du temps : (doc 3 p 369) : il faut au moins 3 semaines pour détecter par des tests
les anticorps anti-VIH.
La lutte contre les virus déclenche souvent les mêmes symptômes : les mêmes qu’un rhume banal (il existe
toutefois des cas d’éruptions cutanées symptomatiques) c’est pour cela que la primo-infection ne permet pas de
diagnostiquer le SIDA.
Concl : Pendant la primo-infection, les symptômes se limitent le plus souvent à ceux d’une maladie virale
bénigne (rhume). Ces symptômes résultent de l’augmentation de la charge virale due à la multiplication des
virus dans leurs cellules hôtes. Le système immunitaire est assez efficace pendant la primo-infection puisque
la charge virale diminue et reste faible pendant la phase suivante dite asymptomatique (p366).
La multiplication du VIH est basée sur une enzyme virale, la transcriptase inverse, qui transcrit l’ARN viral
en ADN dans les cellules infectées. Cet ADN est intégré au génome de la cellule et s’exprime, permettant la
reproduction du virus sous forme de particules virales infectieuses et leur dissémination notamment dans les
organes lymphoïdes (doc p 378)
Quel est le déroulement de la réponse immunitaire ?

Chapitre 2 : Diminution de la charge virale et période asymptomatique.
Deux semaines à quelques mois après la contamination (deuxième partie de la primo-infection), la présence dans le
sang de différents anticorps anti-VIH est décelée, le sujet est dit alors “séropositif pour le VIH”.
en même temps dans le sang du sujet contaminé des lymphocytes T cytotoxiques spécifiques dirigés contre les
cellules infectées par le VIH apparaissent.
Comment les Ac et les LTc provoquent-ils la diminution de la charge virale ?
I. LES ANTICORPS : AGENTS DU MAINTIEN DE L’INTEGRITE DU MILIEU EXTRACELLULAIRE.
Les anticorps sont des protéines d’une famille particulière : les immunoglobulines (p 390/391).
Activité p 391.
Les immunoglobulines sont présentes chez tous les mammifères et sont subdivisées en 5 classes: IgG, IgA, IgM,
IgD et IgE. Elles ont toute une structure de base faite de quatre chaines peptidiques: deux longues (lourdes ) et deux
courtes ( légères ). Elles sont bifonctionnelles avec deux sites réactifs d'attachement aux antigènes a une extrémité
(Fab ) alors que l'autre extrémité ( Fc ) permet la fixation aux cellules ou au complément.
Le type d’immunoglobulines est déterminé par la partie constante de la chaîne
lourde : alpha pour IgA, gamma pour IgG, delta IgD, epsilon IgE, mu IgM.
Un anticorps reconnait plus une structure spatiale que la nature chimique d'une
molécule. La spécificité vis à vis d'une configuration spatiale ( epitope ) est
très grande et distingue plusieurs sites sur une protéine donnée ( un tripeptide
sera reconnu au sein d'une hélice alpha et ne sera pas reconnu en dehors et
vice versa )
A. La spécificité des anticorps.
1) Mise en évidence : le test d’Ouchterlony. TP2
Injection de sérum de lapin à une chèvre : elle produit des Ac-anti lapin
polyclonaux
Test Ouchterlony = immunodiffusion : puits central sérum de chèvre
contenant Ac anti-lapin et dans les puits périphériques sérum d’autres espèces
dont lapin et chèvre.
Les Ac anti-lapin produits par la chèvre se lient spécifiquement aux Ag contenus dans le sérum de lapin. Cela
provoque la formation d’un arc de précipitation : il marque l’emplacement des complexes immuns : faire un dessin.
La diffusion des anticorps dans la gélose nous montre aussi que Les anticorps agissent dans le milieu extracellulaire
(ou milieu intérieur) : ce sont des molécules solubles dans les liquides extracellulaires (sang et lymphe).
2) Le test ELISA.
Modalités générales TP2 : recherche d’un allergène (antigène), la bêta lactoglobuline par un ELISA sandwich
On peut utiliser la spécificité des anticorps pour détecter les antigènes contre lesquels ils réagissent dans
diverses substances à tester.
3) Les tests de dépistage : Cas de la recherche d'anticorps anti-VIH
a) ELISA indirect
Dans le cas du test ELISA appliqué à la recherche d'une éventuelle séropositivité pour le VIH, les protéines de
l'enveloppe et du corps du VIH sont adsorbées en tant qu'antigènes en phase solide dans le puits. Les individus s'ils
sont infectés par le VIH possèdent dans leur sérum des anticorps dirigés contre les épitopes de ces protéines
virales. Ces anticorps sériques contre le VIH peuvent être détectés dans les six semaines qui suivent l'infection.
Les recommandations de fiabilité indiquent qu’il faut attendre le résultat d’un test effectué 3 mois après le rapport
non protégé pour être sûr de son résultat (fenêtre sérologique plus ou moins importante sans qu’il y ait d’Ac anti-
VIH).
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%