JOSSELIN Hélène

1/9
JOSSELIN Hélène
JOUANNO Louis-Marie
07/09/10
Physiologie, Physio cardiaque, Y LESSARD
PHYSYOLOGIE CARDIAQUE
Avant-propos
I- Introduction générale
Revoir le cours de M1 : évolution des concepts, principes de fonctionnement des priorités, dispositifs de
contrôle du SNV.
II- Description anatomo-fonctionnelle
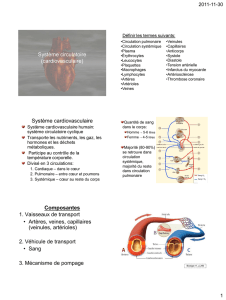
A- Circuit général
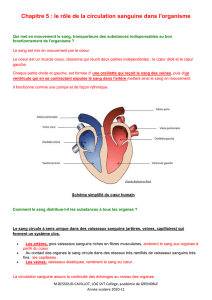
Nous partons du cœur gauche :
Sang arrive à pression quasi nulle de l’oreillette gauche dans le ventricule gauche, à travers la valve
mitrale. Le sang est déjà oxygéné.
Le ventricule élève la pression à 120-150 mmHg.
Le système artériel qui suit fait partie du circuit à haute pression :
Sang éjecté dans l’aorte à travers la sigmoïde aortique, et conduit jusqu’aux organes/tissus selon les
besoins.
Le sang pénètre dans les capillaires, on retourne dans le système à basse pression.
Les capillaires sont perméables à l’eau et molécules diffusibles, donc passage de métabolites et gaz par
diffusion et filtration. Ceci permet l’apport d’éléments indispensables aux cellules et l’élimination des
déchets.
A la sortie des organes le sang est désaturé en O2, il passe dans les veinules-veines et enfin revient
dans l’oreillette droite.
Il passe dans le ventricule droit par la tricuspide et est éjecté dans la circulation pulmonaire par la
sigmoïde pulmonaire à 10-20 mmHg.
Le passage dans les capillaires pulmonaires permet uniquement des échanges gazeux : relargage
de CO2 et oxygénation du sang. Retour à l’oreillette gauche par les 4 veines pulmonaires.

2/9
B- Remarques importantes
1- Artère :
Vaisseau par lequel le sang est évacué du cœur (ventricule)
2- veine :
Vaisseau par lequel le sang revient au cœur.
Il existe 2 types de circulation :
Circulation systémique : sang veineux =sang désaturé en O2, sang artériel=sang oxygéné
Circulation pulmonaire : artère pulmonaire contient du sang désaturé, veines pulmonaires contiennent du
sang oxygéné.
3-myocarde :
Il n’y a pas d’échanges de flux entre oreillettes ou ventricules.
4- Les bronches :
Sont au contact du sang pour leur propre vascularisation et non pour des échanges. Le sang de cette
vascularisation a 2 destinés : -retour à l’oreillette droite par veine cave supérieure –retour à l’oreillette
gauche par les veines pulmonaires (shunt)
5- le foie :
Fonction d’épuration du sang désaturé mais aussi oxygéné arrivant par l’artère hépatique.
6- Le rein :
Le sang oxygéné (rouge) arrive aux glomérules et irrigue les tubules.

3/9
III-Caractéristiques mécano-histologiques du réseau
vasculaire
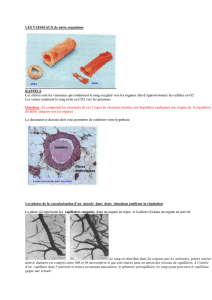
Structure des vaisseaux : fig.2 page 2 poly
Les vaisseaux comportent 3 couches :
- intima : épithélium en contact du sang – fibres élastiques longitudinales
- média : surtout des fibres élastiques circulaires- peu de fibres musculaires lisses
- adventice : fibres longitudinales de collagène
La proportion des éléments peut varier dans les couches, donc les propriétés sont différentes selon ces
proportions.
Soumise à un étirement :
Fibre élastique augmente beaucoup en longueur mais provoque peu de tension
Fibre collagène s’allonge peu mais voit sa tension augmenter très vite.
NB : les artères avec beaucoup de fibres collagènes ont une faible compliance = distancibilité
Plus le diamètre des artères /artérioles diminue, plus la résistance est élevée. Ce phénomène est majoré
par vasoconstriction active. Artères et artérioles sont donc résistives.
Les capillaires ont un endothélium (1 seule couche de cellules et n’ont pas de fibres).
Veinules/veines contiennent beaucoup de fibres musculaires lisses (comme les artérioles) mais
moins de fibres élastique/collagène. Ce sont des vaisseaux capacitifs : capacité à contenir le sang.
A l’entrée des capillaires on trouve des sphincters (anneaux de fibres musculaires lisses) qui se
ferment/s’ouvrent pour permettre les échanges.

4/9
A-Modèles de réseaux vasculaires
Le sang est distribué dans un circuit très ramifié dont les propriétés sont variables. Les calculs
pour comprendre les réseaux sont basés sur des modèles. Ces modèles permettent la schématisation des
réseaux vasculaire, ce sont des réseaux à progression géométrique : fig.3 page 2 poly.
Les formules donnent le nombre de vaisseaux, leur diamètre et leur longueur.
Ex : étage 0 : 1 vaisseau – l’aorte
diamètre -2cm
longueur-59cm
étage 8 : 3milliards de vaisseaux (capillaires)
diamètre-8 µm
longueur-1,5 mm
B- réseaux vasculaires naturels
« Ça colle pas mal » !
Les calculs faits à partir des réseaux schématiques concordent avec les réseaux naturels.
On peut les étudier dans des zones particulières comme la rétine, réseau en 2D.
1- Section de passage
Définition : c’est la somme des vaisseaux d’un même étage, elle augmente continuellement vers
les capillaires puis diminue au retour par les veinules/veines. Schéma page 2 poly
2-Vitesse d’écoulement
Le débit est constant à tous les étages (circuit fermé), il est de 5,2-5,4 L/min. Donc la vitesse est
plus faible dans les artérioles et capillaires.
3-Pression
Séparation brusque entre secteur haute/basse pression au niveau des artérioles, ceci est dû aux
frottements de la paroi des vaisseaux. La pression reste basse dans le circuit pulmonaire.
NB : ramifications naturelles
Séparation des vaisseaux suivant des angles variables qui dépendent du diamètre des vaisseaux
naissants. Ex : gros diamètre-angle petit
Petit diamètre-angle grand

5/9
IV-Irrigation des principaux territoires
A- Rôles du réseau
1- rôle statique de capacité
Les veines vont contenir une grosse partie de la masse sanguine.
2- rôle dynamique de répartition
Se fait en fonction des besoins de l’organisme ; de façon à préserver les zones dont le débit ne doit
pas varier (cerveau) et à irriguer plus ou moins d’autres régions selon les besoins.
B- Caractéristiques circulatoire des régions
Tableau poly page 3
NB : retenir les valeurs comparatives
La consommation d’Oxygène représente les besoins de l’organisme. Le cœur a besoin d’un gros
débit car il consomme beaucoup d’oxygène même au repos.
A l’effort le débit peut être multiplié par 5-6, ex : poumons, mais peut diminuer ex : système
splanchnique (mésentère/foie/rate) . Ce sang sera redirigé vers des zones où les besoins augmentent.
Les glandes endocrines ont une très forte consommation d’oxygène au repos comme à l’effort car
elles synthétisent des hormones en permanence.
Le débit du cerveau ne DOIT pas changer.
 6
7
8
9
6
7
8
9
1
/
9
100%