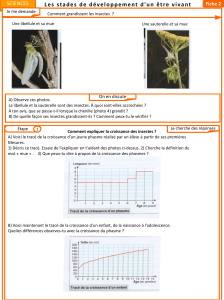
Nutrition et développement post embryonnaire des arthropodes

Nutrition et développement post
embryonnaire des arthropodes autres que
les insectes (aspects morpho anatomiques et
contrôle endocrine).
DEVELOPPEMENT POST EMBRYONNAIRE DES CRUSTACES
A. RAPPELS DE CLASSIFICATION
Voir diapo
Crustacés : 7 paires de pattes et 2 paires d’antennes
Ils comportent un exosquelette rigide comportant jusqu’à 50% de cristaux de calcium (ou sel ?)
B. QUELQUES RAPPELS DE BIOLOGIE DES CRUSTACES
Taxonomie : environ 45 000 espèces
Groupes les plus connus : Branchiopodes, Ostracodes, Copépodes, Cirripèdes, Malacostracés.
Deux paires d’antennes ( exception : deuxième paire rudimentaire chez les isopodes
cloportes)
La structure fondamentale des appendices est de type biramé.
Schéma
Appendice biramé Endopodite (branche interne, locomotion), exopodite (branche
externe, respiration) portée par le basipodite. Parfois il ne reste que l’endopodite.
Animaux marins (majoritairement)
Plusieurs stades larvaires
Les trois premières paires d’appendice servent à la nage chez la larve
Développement de type anamorphique (ou développement dilaté) : on l’oppose au
développement épimorphique
Au moment de l’éclosion, tous les segments qui composent le corps de la larve ne sont
pas formés
Les nouveaux segments s’ajouteront en avant du pygidium (telson) au fur et à mesure
des mues.
Structure des adultes :
Exemple de la langoustine :
Tête = Céphalon : 6 segments, chaque segment comportant des appendices
Thorax = Péréion : pattes ambulatoires et 8 segments
Abdomen = Pléon 6 segments (nombre de segments variable)
Céphalon + Péréion = Céphalothorax

Chez les insectes, le thorax est constitué de trois segments.
Une paire d’appendice par segment
Chez les rémipèdes et les malacostracés, chaque segment (somite) porte un appendice.
C. LES STADES DE DEVELOPPEMENT CHEZ LES CRUSTACES
5 stades de développement. Exemple de la crevette (comporte l’intégralité des stades de
dévelppement) :
Nauplius
Metanauplius
Protozoe
Zoe
Mysis
La larve nauplius est une larve de type planctonique (aquatique errante), sans capacité natatoire
réelle. Pour la crevette, la taille de cette larve est de l’ordre de 0.5mm. elle a une forme ovoide. On
va avoir du mal à distinguer les traces de segmentation. On trouve trois paires d’appendices :
Les antennules (A1)
Les antennes (A2)
Les mandibules
On a également un œil unique, un orifice buccal. L’épistome est la zone de croissance entre les
antennules et la bouche.
Sur les trois paires d’appendices, seules deux paires (A1 A2) sont de type biramé. Elles, à elles seules,
vont assurer un rôle sensoriel, alimentaire et moteur (en gr orienter la direction, elles vont orienter
la direction, vaguement).
Sur la partie caudale, elle comporte deux longues soies caudales qui vont progressivement se
développer en nageoire caudale plus tard.
Durée du stade= 1 jour et demi
Il n’y a pas d’ajout d’appendices sur la larve metanauplius. Cependant, il y a de nombreuses
modifications de l’organisation interne.
On a un ajout de quatre segments (métamères) au niveau de la partie caudale. Elle mesure 0.7mm.
la durée du stade metanauplius est de 1 jour et demi.
Les soies caudales se sont différenciées et donnent place à une furca bifide.
Sur la larve protozoe, on distingue l’ensemble des appendices pairs du céphalon. L’œil nauplien est
toujours fonctionnel (permet la vue) mais on commence à observer les yeux définitifs (on les observe
par transparence à travers le tégument)
on a l’apparition de pattes mâchoires sur les deux segments métamériques apparus au stade
métanauplius (les deux autres portant les appendices du céphalon ).
Différenciation de plus en plus importante de la furca.
Taille : 1.5mm
Durée du stade : 2 jours environ
Le stade zoe dure environ trois jours et la larve fait 2-2.5mm.
Disparition de l’œil nauplien, la paire d’yeux devient fonctionnelle. On a des soies qui apparaissent
sur les appendices. On a l’apparition des pattes mâchoires (Pmx3)
Taille : 2-2.5mm

Durée : 3 jours
Le stade mysis est celui qui ressemble le plus à l’adulte. D’ailleurs, il précède l’état adulte.
C’est le stade de développement le plus long (10-15 jours) où tous les appendices manquant
apparaissent (notamment tous les appendices abdominaux et les pléopodes). Très forte
différenciation de la partie caudale avec la formation progressive de deux uropodes qui, avec le
telson (dernier appendice abdominal), forment la nageoire caudale.
Il y a une fusion des segments thoraciques et de la tête formation de la carapace
céphalothoracique.
Nage orientée
L’apparition de la capacité à se reproduire marque la fin du stade mysis et le début de l’âge adulte.
Le stade nauplius et metanauplius peuvent se dérouler à l’intérieur de l’œuf.
Chez les rimipèdes, on parle d’orthonauplius, de metanauplius (1-2-3 selon l’avancement des mues)
et de post nauplius.
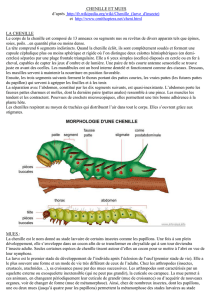
LA MUE ET SON CONTROLE
A. LES MUES
Fonctions de l’exosquelette :
Barrière, protection contre les agressions environnementales
Fixation des muscles dessus
L’exosquelette est rigide donc si la larve grossit, elle va se retrouver bloquée par celui-ci.
La cuticule :
Epicuticules
Exocuticule
Endocuticule
Epiderme
L’ épicuticule a juste une lame chez les crustacés. Elle est constituée de composés appelés cuticuline
(surtout lipides, protéines, glucides)
Les lipides ont une fonction de protection contre la déshydratation.
L’exo et endocuticule forment la procuticule..
Sous l’endocuticule, on a les cellules épidermiques. Ce sont elles qui vont être responsables de la
dégradation de l’ancien exosquelette et de la synthèse du nouvel exosquelette.

A la fin de la période de croissance inter-mue, on a l’apolyse (décollement des cellules de L’épiderme
par rapport à l’endocuticule).
Les cellules de l’épiderme s’agrandissent dans l’espace formé. Elles vont sécréter des enzymes qui
vont pouvoir créer le nouvel exosquelette elles synthétisent le gel de mue ( présence de
protéases, chitinases)
C on a d’abord la mise en place, juste au dessus des cellules épidermiques, de la nouvelle
épicuticule. comme ça, quand le gel de mue arrive sur l’ancienne endocuticule pour la détruire, les
enzymes d’abord inactives présentes dans le gel, s’activent au niveau de l’endocuticule et la digèrent
sans pouvoir attaquer les cellules épidermiques à cause de la nouvelle épicuticule.
Les composés de l’endocuticule sont recyclés en passant à travers la nouvelle épicuticule par
transport et les cellules épidermiques fabriquent une nouvelle endocuticule à partir de ces derniers.
D les lignes d’exuviation (de faiblesse de la cuticule) permettent à l’animal de se dégager de son
ancienne exocuticule par contorsion.
Gonflement de l’animal par absorption d’eau . deux fonctions :
déplisser les tissus
remplit l’espace libéré en attendant que le crustacé grandisse et l’occupe avant
sa prochaine mue.
Durcissement (minéralisation) de la nouvelle cuticule

L’animal va d’abord se débarrasser de la partie antérieure de son ancien exosquelette puis, du pléon
par la suite.
A la fin, l’animal consomme son ancien exosquelette pour récupérer une partie des substances
comme les glycoprotéines qui vont être utilisées pour la consolidation du nouvel exosquelette.
B. LES DIFFERENTES ETAPES DE LA MUE
On va distinguer différentes étapes pour le cycle de la mue (il faut voir ça comme un phénomène
cyclique et non linéaire).
Stade A = Métecdysis
Stade B = Metecdysis
Stade C = Anecdysis
Stade D= Proecdysis
Stade E = Ecdysis (mue)
Stade A : metecdysis : A1 et A2 (2% du temps)
Période qui suit directement la mue. Un jour environ. Tissus remplacés par de l’eau.
Les animaux sont inactifs et ne s’alimentent pas. (consommation des réserves : triglycérides
et glycogène)
L’exosquelette est mou, et non calcifié
Epiderme transparent et peu pigmenté.
Ce qui sépare A1 et A2, ce sont des processus physiologiques qui ont lieu durant cette
journée.
Formation de l’épicuticule et de l’exocuticule par les cellules épidermiques
Stade B : metecdysis : B1 et B2 (8% du temps)
Exosquelette qui durcit progressivement du à un dépôt de calcium et substances minérales
Sécrétion de l’endocuticule
Accroissement de la pigmentation de l’épiderme
Fort accroissement de la synthèse d’ADN et de protéines
Reprise progressive de l’activité locomotrice
Stade C : anecdysis : C1 C2 C3 et C4 (66% du temps) phase de croissance intense (inter mue)
Calcification de l’exosquelette terminée
Activité des animaux maximale et alimentation
Période de croissance des tissus
Exosquelette rigide
Perte d’eau
Forte synthèse de réserves corporelles (triglycérides, glycogène, hexamérides) au niveau de
l’hépato pancréas
Plus l’animal vieillit, plus le stade C va être long.
Stade D= proecdysis. D1 D2 D3 et D4 (24% du temps)
Période avec des changements actifs au niveau morphologique et physiologique (préparation de la
mue)
Activation des cellules de l’épiderme
Analyse, séparation des cellules de l’épiderme et des cellules de l’endocuticule
Début de résorption de l’ancien exosquelette
 6
7
8
9
6
7
8
9
1
/
9
100%