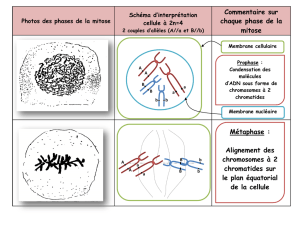
cycle cellulaire - E

Tous droits réservés 2009-2010 © www.objectif-p1.com
C
CY
YC
CL
LE
E
C
CE
EL
LL
LU
UL
LA
AI
IR
RE
E
I
I.
.
I
IN
NT
TR
RO
OD
DU
UC
CT
TI
IO
ON
N
A. Différences entre Eucaryote et Procaryote en ce qui concerne le cycle cellulaire.
Procaryote : Réplication de l’ADN en permanence, donc division très rapide.
Il n’y a donc qu’une seule et même phase.
Eucaryote : Réplication et division sont dans des phases bien distinctes.
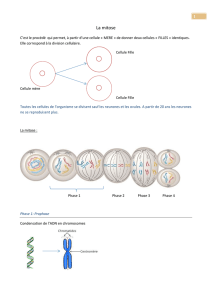
B. Le cycle se divise essentiellement en 2 phases :
Mitose = Phase de division.
Interphase = intervalle entre 2 divisions
G1
Gap 1
intervalle 1 : sépare M et S
S
Synthèse
G2
Gap 2
intervalle 2 : sépare S et M
Et il y à aussi une phase G0 (Gap 0) qui constitue une phase de quiescence.
D. Courbe représentant la quantité d’ADN en fonction des différentes phases.

Tous droits réservés 2009-2010 © www.objectif-p1.com
E. Phase G0.
Durée : Durée qui dépend de la présence de facteur de croissance et des types cellulaires.
Conditions : La phase G0 n’a pas de facteur de croissance.
F. Si la cellule subie un dommage trop important.
Elle sort du cycle, va en apoptose (mort cellulaire programmée)
G. Durée des différentes phases.
G1
9 à 10 h
S
9 à 10 h
G2
4 à 5 h
M
Environ 1h
G0
Variable
G. Fréquence des cycles.
Très variable d’un type cellulaire a un autre
2 exemple :
Les fibroblastes ont 1 mitose par an.
Les hépatocytes ont 2 mitoses par an (1 tout les 6 mois), plus si stimulation.

Tous droits réservés 2009-2010 © www.objectif-p1.com
I
II
I.
.
E
ET
TU
UD
DE
E
D
DE
E
L
L’
’I
IN
NT
TE
ER
RP
PH
HA
AS
SE
E.
.
G1
Lors des 1ère divisions de fécondations G1 est quasi inexistante, la cellule passe de S à M et M à S
très rapidement.
Synthèse :
La cellule synthétise des protéines (qui lui permettent d’assurer son bon
fonctionnement).
Elle synthétise aussi de l’ARN (t, m, r) qui lui permettra d’assurer son accroissement.
Elle synthétise des facteurs de croissance qui permettent une autostimulation afin de
rester dans le cycle.
Elle synthétise les protéines qui vont s’associer a l’ADN pour permettre sa
réplication.
Cette phase est divisée en 2 sous phases séparées.
Sous phase 1 : La phase G1-pm qui nécessite des facteurs de croissance
Sous phase 2 : La phase G1-ps qui est indépendante de la présence de facteurs de
croissance.
Séparateur : Le point R , point de non retour a partir duquel la cellule n’a plus besoin de
facteurs extérieurs.
G1-pm
1) Durée : 3 heures
2) Fonction : Temps nécessaire a la modification des lamines nucléaires (ajout d’une queue
lipidique d’ancrage) avant acheminement.
3) Substances synthétisées : Elle synthétise en permanence des facteurs de croissance.
4) Action sur le Cytosquelette : Réorganisation et formation du cytosquelette.
G1-ps
1) Phase indépendante de la présence de facteurs de croissance.
2) Accroissement de la taille de la cellule en vue de la duplication de l’ADN.
3) Les protéines s’accumulent de le cytoplasme.
4) Phase de contrôle de l’ADN (réparation), présence d’un check point.

Tous droits réservés 2009-2010 © www.objectif-p1.com
S
Fonction : Phase de duplication de l’ADN.
Durée : Variable, mais en générale 9 à 10 heures.
Pour les cellules embryonnaires : Quelques minutes
Pour les cellules somatique : quelques heures (9 ou 10)
Durant cette phase, la cellules dédouble 2 choses :
La cellule dédouble son centrosome afin de permettre la formation du fuseau mitotique
durant la mitose.
La cellule dédouble son ADN afin de permettre une répartition égale de son matériel
génétique en deux cellule fille.
En cas d’erreur durant cette phase, il existe un mécanisme de correction des erreurs de copie :
la correction sur épreuve.
Si les erreurs sont trop importante la cellule sort du cycle et se met en apoptose.
G2
Cette phase sépare : La phase de duplication (S) de la phase de division (M)
Durée : 4 à 5 heures.
Cette phase a pour fonction de vérifier la bonne réplication de l’ADN, elle vérifie et corrige si
nécessaire les erreurs.
Elle s’assure que toute les structures permettant la mitose (taille de la cellule, organites, ADN,
etc.) sont en place.
La synthèse d’ARN et de protéine continue.
G0
Cette phase est dite de repos ou de quiescence
Durée : très variable d’un type cellulaire a un autre, de quelques heures, a plusieurs
années.
Au microscope elle ressemble a une cellule en G1 (difficile a distinguer) pourtant elle a un
métabolisme différent. En effet elle produit (synthétise) beaucoup moins.
Si elle est stimulée avec des facteurs de croissance, elle retourne dans le cycle cellulaire, en G1.
Ex : en cas de biopsie du foie, les hépatocytes en G0 vont s’activer et se diviser quasi
quotidiennement.
Protéine qui n’est pas présente durant cette phase : La cycline.

Tous droits réservés 2009-2010 © www.objectif-p1.com
I
II
II
I.
.
L
LO
OG
GI
IQ
QU
UE
E
D
DE
E
C
CY
YC
CL
LE
E.
.
A. Intérêt de connaître le déroulement de ce cycle.
Intérêt thérapeutique, la réponse diffèrent d’une phase a une autre.
B. 3 expériences prouvent que l’ordre de ce cycle est obligatoire.
G2 + M
G1 + S
G2 + S
Des facteurs dans la cellule en M
induisent le passage en phase de
division chez la cellule en G2
Des facteurs dans la cellule en S
induisent le passage en phase de
réplication chez la cellule en G1
Pour empêcher la tétraploïdie,
une cellule en G2 ne peut pas
repassé en S (facteur inhibiteurs)
Phases
Réplication +
correction sur
Epreuve
ADN ARN
ARN Protéine
Réparation de l’ADN
G1
NON
OUI
OUI
OUI
S
OUI
OUI
OUI
NON
G2
NON
OUI
OUI
OUI
M
NON
NON
+/- NON !!!
NON
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%