TP 2 - Free
publicité

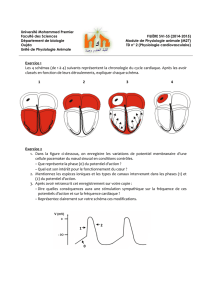
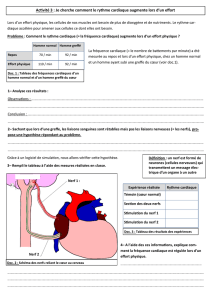
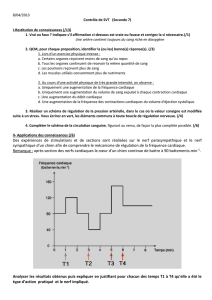
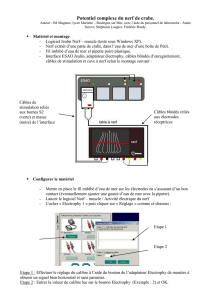
Destrez Guillaume Chateigner Aurélien A2 TP2 : Activité électrique du nerf TP2 : Activité électrique du nerf 1ère partie : Mesure à l’oscilloscope cathodique 2ème partie : Mesure en EXAO Introduction : Ce TP a pour but l’étude des caractéristiques du potentiel d’action du nerf sciatique de la grenouille, en utilisant un oscilloscope cathodique dans un premier temps, puis un logiciel EXAO dans un second temps. Nous chercherons, à l’aide de l’EXAO et de l’oscilloscope, à considérer le décours du potentiel d’action et les caractéristiques de la réponse nerveuse, à estimer la vitesse de conduction du nerf sciatique de la grenouille, à approcher les variations d’excitabilité du nerf, à nous assurer que la conduction du nerf se fait bien dans les deux sens, à expliquer l’effet ON-OFF, et enfin à nous rendre compte des effets d’une lésion et d’un anesthésiant. Le potentiel d’action est la réponse caractéristique entrainée par une excitation suffisante d’une fibre nerveuse. Il est la variation caractéristique du potentiel de membrane. C’est un phénomène électrique qui présente deux propriétés remarquables : lorsqu’il se développe, il le fait d’une manière « tout ou rien », et lorsqu’il est émis en un point, il se propage sans atténuation. 1. Résultats a. Observation de la réponse électrique globale Nous plaçons les électrodes en position trois et quatre. La stimulation est faite pendant une faible durée (0,1 ms) et l’intensité de stimulation est augmentée jusqu’à obtenir une réponse électrique globale maximale. C’est au niveau des électrodes de stimulation que la stimulation se fait et entre les électrodes réceptrices que le potentiel d’action et recueillie. Courbe 1 : Voltage de stimulation : 510 mV Amplitude du potentiel d’action : 120 mV Durée du potentiel d’action : 0,2 ms Temps de latence : 1,2 ms 1 Destrez Guillaume Chateigner Aurélien A2 b. Caractéristiques de la réponse nerveuse TP2 : Activité électrique du nerf La courbe 2 nous permet d’illustrer les quatre différentes réponses que l’on peut obtenir. De zéro à 190mV, il n'y a pas de réponse. À 230 mV, on obtient une courbe minimale. À 240 mV, on obtient une réponse normale. À 510 mV, on obtient une réponse maximale. c. Conduction dans les deux sens En inversant le sens de stimulation, c'est-à-dire en stimulant du côté distal, la réponse nerveuse observée et d'une amplitude plus faible que dans l'autre sens. Cette observation nous permet d'affirmer qu'il y a bien conduction dans les deux sens, définissant le nerf sciatique comme un nerf moteur. En effet, si dans le sens normal on obtient une réponse de 23 mV, dans le sens inverse on obtient une réponse de 0,3 mV. d. Détermination de la vitesse de conduction Dans cette expérience, nous étudions la différence entre la réponse entre deux positions d'électrodes différentes. Nous observons alors une plus faible amplitude de la réponse lorsque les électrodes sont en position quatre et cinq, que lorsqu'elles sont en position trois et quatre. Sur la courbe, on observe un décalage entre les deux temps de latence, ce qui décale les amplitudes. On mesure alors grâce au delta X-Y le temps de conduction entre les deux électrodes. On obtient 0,24 ms. On a mesuré une distance de 8 mm entre les deux électrodes, et grâce au temps de conduction entre les deux électrodes on peut déterminer la vitesse de conduction : v = d/T = 8.10-3/0,24.10-3 = 33,33m.s-1 Calcul d'erreur : Δv/v = Δd/d+ ΔT/T = 1.10-3/8.10-3+0,01.10-3/0,24.10-3 = 0,17 Δv = vx0,17 = 33,33x0,17 = 5,56 v = 33,33m.s-1 ±5,56 e. Etude de la relation entre intensité et durée du stimulus Lorsque la durée d'un choc tend vers l'infini, l'intensité minimale nécessaire pour déclencher une réponse liminaire est appelée rhéobase. Sur la courbe, la rhéobase est atteinte à 0,8 ms pour un voltage de 50 mV. La durée nécessaire pour déclencher une réponse liminaire pour une intensité de stimulation égale à deux fois l'intensité rhéobasique est appelé la chronaxie. Sur la courbe, la chronaxie vaut 0,2 ms. La courbe est divisée en trois parties. La première partie se trouve sous la courbe, la deuxième partie est la courbe elle-même, la dernière est la zone au-dessus de la courbe. 2 Destrez Guillaume Chateigner Aurélien A2 f. Etude des périodes réfractaires TP2 : Activité électrique du nerf i. par la méthode des seuils Lors de cette expérience nous effectuons deux stimulations successives d’une durée de 0.1ms séparées par un délai important en choisissant un réglage supraliminaire pour la première stimulation et liminaire pour la seconde. On réduit ensuite le délai qui sépare les deux stimulations tout en veillant à ce que la deuxième soit toujours liminaire. En effet nous nous intéressons ici seulement aux fibres les plus excitables du nerf en cherchant leur seuil d’excitabilité. Sur la courbe représentative de l’évolution du voltage liminaire en fonction du temps entre deux chocs on observe deux phases : -Une première phase avec un délai inférieur à 0.5 ms qui correspond a une réponse absente sur l’oscilloscope, même si l’intensité de stimulation est augmentée. -Une deuxième phase qui correspond a un délai supérieur à 0.5 ms, avec des intensités importantes au début, qui diminue au fur et a mesure que l’on augmente le délai. ii. Par la méthode des amplitudes : Ici nous effectuons aussi deux stimulations successives de 0.1 ms séparées par un délai important, mais en choisissant des réglages d’amplitudes maximale pour les deux chocs. Ce sont donc toutes les fibres qui nous intéressent ici. On réduit ensuite progressivement ce délai pour obtenir une chute progressive de l’amplitude de la 2ème réponse jusqu’à sa disparition. On obtient les résultats suivants : Délai entre 2 stimulations (ms) Amplitude de la réponse (mV) 1 2 3 5 0 2.5 4 5 % du potentiel d’action maximal (4.9mV) 0 50 80 100 3 Destrez Guillaume Chateigner Aurélien A2 g. Effet on-off TP2 : Activité électrique du nerf On applique sur le nerf une stimulation supraliminaire de longue durée (≥5ms). On observe une première réponse d’amplitude maximale. On devrait normalement obtenir deux réponses successives, une A1 au début de la stimulation et une A2 à la fin de la stimulation avec A1 ayant une amplitude supérieure à A2. Nous aurions du donner une stimulation de plus longue durée ou changer la fréquence pour voir sur une plus longue durée… h. Etude de l’essai de deux chocs sous liminaires On effectue deux chocs sous liminaires séparés par un délai assez long. On n’observe aucune réponse. En séparant les deux chocs de seulement 0.2ms on observe une réponse supra liminaire. i. Observation des effets d’un anesthésique Nous plaçons un coton imprégné d’éther dans la cuve a nerf et on stimule le nerf toutes les 30 secondes afin d’obtenir une réponse d’amplitude maximale. On observe une amplitude de la réponse qui diminue rapidement. Une fois que la réponse a atteint 10% de la réponse maximale on enlève le coton d’éther et on stimule toutes les 30 secondes : On observe aux environs de 2min 30 après avoir enlève le coton, que le nerf redonne une réponse de faible amplitude jusqu'à redonner une réponse maximale. Cependant suite a une erreur d’enregistrement des fichiers a cause d’une mauvaise saisi de nom de fichier nous avons du prendre les courbes d’un autre groupe et donc n’avons pas notés tous les temps. j. Effet d'une lésion irréversible sur le nerf Cette expérience consiste à pincer le nerf entre les électrodes 4 et 5 et d’étudier sa réponse après une stimulation d’amplitude maximale. On obtient une réponse monophasique (on a qu’une phase de la réponse : la phase positive). 4 Destrez Guillaume Chateigner Aurélien A2 2. Interprétation TP2 : Activité électrique du nerf a. Observation de la réponse électrique globale La courbe 1 obtenue peut être divisé en plusieurs parties. La première partie est le temps de latence, c’est le temps qui sépare l’arrivée de la stimulation aux électrodes de mesures de l’arrivée aux électrodes de stimulation. C’est durant ce temps que l’on observe un artefact des stimulations qui se caractérisent par une variation du potentiel d’action. Il correspond à la réponse passive du nerf et est uniquement physique. La deuxième est la phase au cours de laquelle l’onde de négativité atteint la première électrode réceptrice, et inverse les charges, chargeant le nerf négativement à l’extérieur. La troisième est la phase au cours de laquelle l’onde de négativités quitte la première électrode réceptrice, faisant revenir le potentiel d’action à sa valeur d’origine. Ces deux dernières phases correspondent donc au passage de l’onde de négativité sur la première électrode réceptrice, elles forment donc la réponse bi phasique qui est globalement négative. La quatrième est la phase au cours de laquelle l’onde de négativité atteint la deuxième électrode réceptrice, inversant les charges à la surface du nerf. Au niveau de la deuxième électrode réceptrice le potentiel devient positif et donc la différence de potentiel est positive. La cinquième est la phase au cours de laquelle l’onde de négativité quitte la deuxième électrode réceptrice, c’est un retour à l’équilibre. Le potentiel de la deuxième électrode réceptrice revient à sa valeur, la différence de potentiel est alors égale à zéro. La quatrième et la cinquième phase marquent donc le passage de l’onde de négativité sur la deuxième électrode réceptrice, elles forment donc la réponse bi phasique qui est globalement positive. On peut alors déterminer sur la réponse bi phasique du nerf 2 phases quasiment identiques mais opposées de par leurs valeurs. La deuxième partie de la réponse n’est pas totalement identique à la première car l’onde recouvre encore la première électrode réceptrice lorsqu’elle atteint la deuxième. b. Caractéristiques de la réponse nerveuse Lorsque le voltage de stimulation est insuffisant pour déclenche une réponse globale du nerf aucune fibre nerveuse n’est recrutée. C’est ce que l’on obtient entre zéro et 190 mV. C’est ce que l’on appelle une réponse infra liminaire. 5 Destrez Guillaume TP2 : Activité électrique du nerf Chateigner Aurélien A2 Lorsque le voltage de stimulation atteint la valeur minimale permettant d’obtenir une réponse, les fibres nerveuses les plus excitables sont recrutées. C’est ce que l’on obtient pour une valeur de 230 mV. C’est ce que l’on appelle une réponse liminaire. Lorsque le voltage de stimulation est suffisant pour stimuler une bonne partie des fibres nerveuses, on obtient une réponse. C’est ce qu’il se passe pour une valeur de 240 mV. C’est ce que l’on appelle une réponse supra liminaire. Lorsque le voltage de stimulation et maximal, et permet de stimuler toutes les fibres nerveuses, on obtient la réponse maximale. C’est ce qu’il se passe pour une valeur de 510 mV. C’est ce que l’on appelle une réponse supra liminaire maximale. L'amplitude de la réponse varie en fonction de l'intensité de stimulation. En effet plus on augmente l'intensité de stimulation plus l'amplitude augmente. Ces stimulations sont dites supra liminaires. Une stimulation maximale entraîne une réponse maximale. Une stimulation sous liminaires ne déclenche aucune réponse, car même les fibres les plus excitables ne peuvent atteindre leur potentiel d'excitabilité. Le potentiel d'excitabilité est le seuil à partir duquel une fibre qui reçoit une stimulation effectue une contraction. Dans le cas d'une stimulation liminaire on peut observer une réponse de faible amplitude. En effet, seules les fibres les plus excitables sont stimulées. On atteint le potentiel critique seulement au bout d'un certain temps, puisque il y a un délai de quelques millisecondes, appelé temps de latence, entre l'excitation des fibres musculaires, et le développement d'une tension. Dans le cas d'une stimulation supra liminaire on peut observer une augmentation de l'amplitude de la réponse et une diminution du temps de latence. Le temps de latence est réduit, d’autan que le potentiel critique est atteint plus rapidement que pour une stimulation liminaire. Avec l'augmentation de l'intensité le recrutement est de plus en plus important, provoquant une augmentation de l'amplitude de la réponse. Dans le cas d'une stimulation supra liminaire maximale, l'amplitude de la réponse est maximale et le temps de latence est minimal. En effet, le potentiel critique est atteint très rapidement, réduisant de ce fait le temps de latence. Toutes les fibres sont alors recrutées, ce qui explique que l'amplitude soit maximale. On peut ainsi à en conclure que le temps de latence globale est décomposé en deux facteurs, qui sont le temps de conduction, indépendant de la vitesse, et le temps de réponse de la fibre nerveuse, variant avec l'intensité de stimulation. On peut expliquer la relation entre l'intensité de stimulation et le temps de réponse de la fibre nerveuse en mettant en lumière le fait que c'est le temps de chargement de la capacité membranaire qui est le principal facteur. En effet il diminue lorsque la stimulation augmente. De plus un nerf étant composé de nombreuses fibres nerveuses, l'augmentation de l'intensité de stimulation stimulant chaque fibre diminue le temps de latence globale du nerf. Lorsque toutes les fibres sont recrutées, l'amplitude devient maximale, en réponse à une stimulation maximale. 6 Destrez Guillaume Chateigner Aurélien A2 c. Conduction dans les deux sens TP2 : Activité électrique du nerf On peut expliquer ce résultat en mettant en lumière le fait que seuls quelques axones du nerf courent sur toute sa longueur. Les autres axones servent à former les collatérales, et ne peuvent donc conduire d'informations jusqu'aux genoux. La propagation du potentiel d'action commence à partir du cône d'émergence, à la base du corps cellulaire du neurone (ou le péricaryon) qui fait la sommation des potentiels gradués provenant des synapses situées le long des dendrites et sur le corps cellulaire. La conduction ne se fait normalement que dans un sens, cependant, le prélèvement de notre expérience sectionne les somas, rendant l'orientation fonctionnelle inefficace. d. Détermination de la vitesse de conduction On peut expliquer ce résultat, comme précédemment, par le fait que seuls quelques axones du nerf courent sur toute sa longueur. Nous avons donc plus d'axones jusqu'à la position quatre que jusqu'à la position cinq. Donc, moins il y a d'axones, moins la réponse est grande. Habituellement, on considère la valeur théorique de la vitesse d'un nerf suivante : 30 m.s-1. Notre résultat de 33,33 m.s-1 est proche de cette valeur, et l'on peut le considérer comme acceptable, puisque la valeur de 30 entre dans notre fourchette d'erreur. L'erreur faite peut-être due aux conditions expérimentales telles que la pression et la température, d'éventuelles lésions du nerf, où une erreur matérielle due à une dégradation ou un vieillissement. e. Etude de la relation entre intensité et durée du stimulus La durée de passage du courant est un facteur important de l'intensité du courant. La chronaxie est l'index de l'excitabilité relative des différents tissus. Plus la chronaxie est faible, plus les tissus sont excitables. La première partie de la courbe représente les stimulations sous liminaire, en dessous de la durée minimale de 0,1 ms, peu importe la stimulation, il n'y a pas de réponse. Il faut donc une durée minimale de 0,1 ms pour déclencher une réponse. La deuxième partie représente les stimulations liminaires. De 0,1 ms à 0,8 ms, l'intensité de stimulation minimale décroît, et atteint sa valeur minimale de 50 mV à 0,8 ms. Cette valeur minimale de 50 mV est la rhéobase. La dernière partie correspond aux stimulations supra liminaires et supra liminaires maximales. On peut conclure pour cette courbe que la grenouille peut exécuter des mouvements rapides, étant donné que la chronaxie est de seulement 0,2 ms. Cette courbe permet de plus de déterminer les caractéristiques des chocs de stimulation efficaces : en effet, il suffit de se référer à cette courbe pour savoir quelle stimulation envoyer suivant la réponse attendue. 7 Destrez Guillaume Chateigner Aurélien A2 f. Etude des périodes réfractaires TP2 : Activité électrique du nerf i. Par la méthode des seuils (En étudiant les fibres les plus excitables) : On observe que lorsque le délai est inférieur a 0.5ms il n’y a pas de réponse (pour le deuxième choc : le choc exploratoire) quelque soit l’intensité donnée : nous somme dans la période réfractaire absolue des fibres nerveuses dont l’excitabilité est nulle pour le deuxième choc. En effet il existe une période appelée période réfractaire durant laquelle le déclenchement d’un second potentiel d’action est impossible. Quand le choc est donné dans un délai supérieur a 0.5ms on observe une réponse liminaire sous condition d’augmenter progressivement l’amplitude de stimulation au fur et a mesure que l’on diminue le délai séparant les deux stimulations. Le deuxième choc intervient alors au cours de la période appelée période réfractaire relative des fibres nerveuses les moins excitables. En effet durant cette période les fibres déjà excitées par le premier choc sont de nouveau suscitée pour un deuxième choc mais le deuxième choc étant situé dans la période réfractaire relative, le seuil d’excitabilité des fibres augmente et donc elles réagissent beaucoup moins. On parle ici d’hypoexcitabilité, il faut donc augmenter l’intensité de stimulation. Donc plus le délai sera court, plus les fibres les plus excitables seront en hypoexcitabilité et plus il faudra augmenter l’intensité de stimulation. On observe après un délai de 4 ms, une intensité nécessaire constante, nous ne somme plus en période réfractaire. ii. Par la méthode des amplitudes (En étudiant toute les fibres) : Quand les deux chocs sont séparés de plus de 5 ms on obtient 2 potentiels d’action de même amplitude : nous sommes hors période réfractaire. Les fibres nerveuses ont retrouvées une excitabilité normale : elles peuvent donc toutes être recrutées pour la deuxième stimulation. Lorsqu’on diminue le délai on observe une diminution de l’amplitude. La période réfractaire des fibres les plus excitables est courte, donc ces fibres peuvent être recrutées pour la seconde stimulation, tandis que les fibres les moins excitables sont en hypoexcitabilité. Il y a donc de moins en moins de fibres recrutées en fonction que le délai diminue, on parle de période réfractaire relative. Quand le délai est inférieur a 2 ms il n’y a plus de réponse pour la seconde stimulation, les fibres n’ont plus le temps de retourner a leur excitabilité de base, nous sommes en période réfractaire absolue (Cependant cette méthode a une certaine incertitude au niveau de la période réfractaire absolue). 8 Destrez Guillaume TP2 : Activité électrique du nerf Chateigner Aurélien A2 Les méthodes des seuils et des amplitudes permettent donc de voir l’hétérogénéité des fibres qui composent les nerfs. Elles nous ont permit de différencier les fibres selon plusieurs seuils d’excitabilité. Elles nous ont aussi permis d’étudier les différentes périodes de réfraction et leur utilités : la période de réfraction absolue qui correspond a un délai trop court entre deux stimulations et qui n’entraîne aucune réponse, une période réfractaire relative qui correspond a des réponses de faible amplitudes et une période hors réfractaire qui correspond a des réponses d’amplitude maximale. g. Effet on-off La courbe obtenu est fausse nous allons donc décrire des donnée théoriques. La première devrait servir à l’établissement du courant en prenant la membrane pour un circuit électrique fermé, ce qui correspondrait à l’effet ON. On observerait ensuite à la fin de la stimulation un artefact qui provoquerait l’annulation du courant et serait donc notre effet OFF. La capacité membranaire qui se charge (plus ou moins en fonction de la durée de la stimulation, ici 5 ms, permet une charge complète normalement) provoque une stimulation qui devrait produire une deuxième réponse de plus faible amplitude. Le deuxième temps de latence est normalement plus grand que le premier. En effet l’influx partant de la deuxième électrode stimulatrice (cathode) il va jusqu'à la première électrode réceptrice, le temps de latence est donc composé du temps de réponse du nerf et du temps de conduction du message. Lors de l’ouverture du circuit c’est la première électrode qui correspond à la cathode donc la distance cathode/électrode réceptrice est plus grande. La propagation du flux dure plus longtemps ce qui explique l’augmentation du temps de latence pour la deuxième réponse, le temps de réponse du nerf ne variant pas. La durée plus longue de ce temps de latence est aussi due à la période réfractaire. La diminution d’amplitude qui devrait être observée devrait être dû au fait que toutes les fibres nerveuses n’ont pas récupérés leur excitabilité ce sont donc les plus excitable qui répondent. h. Etude de l’effet de deux chocs sous liminaires On n’observe aucune réponse visible car le pré potentiel généré n’atteint pas le potentiel critique pour les deux stimulations. En réduisant le délai on fini par avoir une réponse supraliminaire a un délai de 0.5ms. On obtient une réponse car la deuxième stimulation intervient avant que le pré potentiel de la première stimulation n’ait eu le temps de revenir à son potentiel de repos, la capacité membranaire n’étant pas totalement déchargée, le potentiel résiduel du 1er choc et le pré potentiel du 2nd s’additionne. Le cumul des deux charges est donc suffisant pour activer toutes les fibres et donc provoquer une réponse supraliminaire. 9 Destrez Guillaume Chateigner Aurélien A2 i. Observation des effets d’un anesthésique TP2 : Activité électrique du nerf La diminution d’amplitude observée est due aux effets de l’éther. En effet la membrane que l’on considère comme un condensateur métabolique chargé est formé principalement de lipides et de protéines, on parle de bicouche phospholipidique. Hors l’éther est un solvant des lipides et donc permet de désorganiser la membrane des nerfs. Cette désorganisation va entraîner une chute de la capacité de la membrane, l’amplitude des courants locaux va donc diminuer ce qui va entraîner une incapacité croissante de l’activité des canaux ioniques. L’influx nerveux sera de moins en moins transmis, on parle d’anesthésie. La charge de la capacité membranaire devient inférieure au seuil d’excitabilité, il n’y a plus de réponse de la part du nerf, on parle d’anesthésie totale. Quand on enlève le coton d’éther on observe une remonté de l’amplitude de la réponse : on observe donc un effet réversible de l’éther sur la membrane du nerf, la bicouche lipidique se réorganise car elle est malléable et va donc retrouver son excitabilité de départ. j. Effet d'une lésion irréversible sur le nerf Nous obtenons une réponse monophasique : en effet ayant effectué une lésion entre les électrodes 4 et 5 l’onde négative arrive à atteindre l’électrode 4 mais est arrêté avant l’électrode 5 a cause de la lésion. Cette dernière électrode restant toujours au potentiel de référence (0) on ne récupère bien que la phase positive de la réponse. Le phénomène du 1) est donc vérifié, la mesure de la réponse du nerf se fait bien par une différence de potentiel car seule la première électrode est active. 10 Destrez Guillaume Chateigner Aurélien A2 Conclusion TP2 : Activité électrique du nerf Ces deux TP nous ont permis d’observer l’activité électrique du nerf sciatique d’une grenouille à travers deux sources : l’EXAO et un oscilloscope. Nous avons pu mettre en évidence les différences biophasique du nerf puis les caractéristiques de la réponse nerveuse, à savoir ses variations d’amplitude et de temps de latence en fonction de l’intensité de stimulation appliquée. De plus, nous avons constaté que la conduction se faisait dans les deux sens in vitro, ce qui n’arrive pas in vivo et nous avons déterminé cette vitesse de conduction. Nous avons aussi étudié ses périodes réfractaires par deux méthodes faisant intervenir différents types de fibres nerveuses. Enfin nous avons étudié l’effet ON/OFF puis l’effet d’un anesthésique et d’une lésion sur le nerf. Nous pouvons dire que ces différents phénomènes s’appliquent à tous les vertébrés et donc aussi à l’homme, seulement les effets de l’éther sont dangereux, pour une longue ou trop forte exposition il peut détériorer les membranes lipidiques irréversiblement. 11