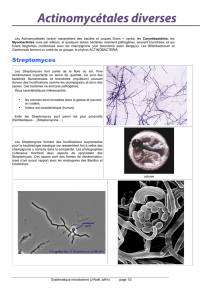
Utilisation de souches de bactéries lactiques

1
!
!
Utilisation de souches de bactéries
lactiques recombinantes en santé
!
!
!
!
MEMOIRE BIBLIOGRAPHIQUE
DU MASTER Sciences et Technologies du Vivant et de l’Environnement
« Nutrition et santé»
Institut National de la Recherche Agronomique
o Adresse : Domaine de Vilvert, 78350 Jouy-en-Josas
o Equipe : ProbiHote, Luis Bermudez et Philippe Langella
o Unité : Microbiologie de l’Alimentation au service de la Santé (MICALIS)
L'Institut des sciences et industries du vivant et de l'environnement, AgroParisTech
Master : Nutrition Santé
Enseignant référent : Anne Marie Davila
Etudiant : Thomas LARCHER, 2ème année de master

2
REMERCIEMENTS
Je remercie Philippe Langella et Luis Bermudez pour m’avoir permis de réaliser ce mémoire
au sein de l’unité d’écologie et physiologie du système digestif à l’INRA de Jouy en Josas.
Je remercie tout particulièrement Luis Bermudez et Anne Judith Waligora-Dupriet pour leurs
disponibilités et leurs conseils.

3
SOMMAIRE
!"!!#$%&'()$*&#+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+,!
-"!./+0#1+'23*1$&*%4+5*)%&6*4##4+""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+,!
7"!80$(%4+'41+9%&6*&$*:(41+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+;!
<"!=%&6*&$*:(41+4$+5>)0#*154+'4+?2*55(#*$>"+""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+@!
!!"!A41+60)$>%*41+?0)$*:(41+B>#>$*:(454#$+5&'*C*>41+""""""""""""""""""""""""""""""""""""""""""""""""""""+D/!
-"!7AEF+G4)$4(%+'20#$*BH#41+4$+'4+90$3&BH#41+""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+DD!
7"!7AEF+9%&5&$4(%+'4+?0+%>9+0#$*I*#C?0550$&*%4+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""+DJ!
<"!7AEF+4$+?20??4%B*4+""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+DK!
L"!7AEF+4$+1&#+0)$*&#+0#$*&MN'0#$4+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+DK!
O"!7AEF+4$+?2&6>1*$>+""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+D.!
P"!A41+6*C*'&60)$>%*41+B>#>$*:(454#$+5&'*C*>41Q+(#4+0G0#)>4+G4%1+'4+#&(G4??41+
$3>%09*41+R+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+D,!
"#!$%!&'(%!)&*+,*-,./%!0%!1*/23%1!14/546%1!0%!+,7,0*+42-8&,%1!####################################################!"9!
:#!!"#"$%&'()*+",-!68;8-,./%<%;-!<*0,7,8!)*/&!08(,5&%&!0%1!)&*-8,;%1!4;-,=
,;7(4<<4-*,&%1!########################################################################################################################################################!">!
E"!<&#)?(1*+4$+94%194)$*G41+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+D@!
!!!"!7*6?*&B%093*4+"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""+S/!

4
SOMMAIRE DES FIGURES

5
LEXIQUE
Ag
Antigène
ANSES
Agence National de Sécurité Sanitaire
BLGM
Bactérie Lactique Génétiquement Modifié
CAT
Catalase
CD
Maladie de Crohn
CPA
Cellule(s) présentatrice(s) d'antigène(s)
DC
Cellule(s) Dendritique(s)
FAO
Food and Agriculture Organization
GALT
Gut Associated Lymphoïd Tissu
GC
Guanine - cytosine
GRAS
Generally Recognized As Safe
hIL-10
Cytokine humaine 10
IFN
Interféron
IL
Interleukine
LAB
Bactérie Lactique (Acid Lactic Bacteria)
LB
Lymphocyte B
MICI
Maladie Inflammatoire Chronique de L'intestin
NDR
Nod Like Receptor
OGM
Organisme génétiquement modifié
PPR
Patterns Recognition Receptor
ROS
Espèces réactives de l'oxygène
SOD
Superoxyde dismutase
TLR
Toll Like Receptor
TNBS
2,4,6-acide trinitrobenzene sulfonic
UC
Rectocolite hémorragique
WHO
World Health Organization
!
!
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
1
/
34
100%