Immunologie : Exercice Corrigé sur la Production d'Anticorps
Telechargé par
Ramdan Chikh

Grâce à cet exercice corrigé de SVT, découvre ce cours complet niveau lycée sur la
notion d'immunologie.
Dans le cadre de ta préparation bac scientifique, ton prof de soutien scolaire en ligne de SVT t'explique
comment on peut rédiger les différentes questions en se souvenant qu'il faut toujours partir des
observations donc des données documentaires.
Examen corrigé sur les conditions de la production d'anticorps

I) a) Le lapin A qui n’a reçu aucune anatoxine meurt systématiquement quelle que soit la toxine injectée :
son système immunitaire ne peut lutter contre ces maladies qui évoluent plus vite que la mise en place de
la réaction immunitaire adaptative.
Le lapin B qui a reçu une injection d’AT (= anatoxine tétanique) seule survit à la TT (= toxine tétanique)
mais pas à la TD (= toxine diphtérique), preuve que la réaction immunitaire qu’il a développée est
spécifique du tétanos : c’est la réaction immunitaire adaptative.
b) L’expérience C montre que par la seule injection du sérum du lapin B au lapin C, celui ci est protégé
contre le seule tétanos mais pas contre la diphtérie (comme le lapin B), preuve que le sérum contient les
effecteurs de la réaction immunitaire spécifique. Or on sait que le sérum ne contient que des
molécules:ces effecteurs sont donc des protéines : les anticorps (Ac). L’expérience D montre que le lapin
perd l’immunité sérique de C lorsque le sérum de ce dernier circule, avant son injection, sur une poudre
imprégnée d’AT : celle-ci retient donc les Ac sériques anti-tétanos, preuve que ces derniers sont
spécifiques de la bactérie du tétanos. Enfin l’expérience E montre que si le sérum de C traverse la poudre
dépourvue d’AT, le lapin E conserve l’immunité anti-tétanique : la spécificité des Ac à l’AT se révèle donc
dans la fixation des Ag à leurs Ac spécifiques. Il y a ainsi formation d’un complexe immun par
complémentarité de forme 3D entre les ag et leurs Ac spécifiques.
c) L’électrophorégramme de B montre un pic plus élevé de gamma-globulines que chez A. Or B seul a
reçu de l’AT donc produit des Ac spécifiques de l’AT que l’on peut ainsi assimiler aux gamma-globulines.
Ce pic constaté permet de pratiquer la sérothérapie chez un sujet en manque d’Ac, mais aussi de repérer
sa séropositivité pour une maladie, c’est-à-dire la fabrication d’Ac spécifique de cette maladie.
d) Le lot A montre que la présence de moelle osseuse permet la production des LT et des LB. Mais le lot B
indique que, si la moelle osseuse suffit à produire des LB, le thymus semble nécessaire à la production
des LT en plus de la moelle osseuse. Enfin, le lot C indique que la seule présence du thymus ne permet
de produire aucun L. Preuve que la moelle osseuse peut être considérée comme le lieu de naissance des
deux types de L. mais le thymus permet d’obtenir des LT fonctionnels alors les LB deviennent fonctionnels
dans la seule moelle osseuse.
e) L’agglutination témoigne de la formation d’un complexe immun (Ag/Ac) donc de la production d’Ac
spécifiques de l’Ag (= pneumocoques). Le lot C montre que sans L, il n’y a aucune agglutination donc
aucune production d’Ac dont on sait qu’ils proviennent des plasmocytes issus de la différenciation des LB.
Avec les seuls LB présents dans le lot B, l’agglutination n’a pas lieu non plus alors que pourtant, on sait
que les LB spécifiques des pneumocoques sont responsables de la production des Ac après leur
différenciation en plasmocytes. Enfin, avec le lot A, les complexes immuns sont nets preuve que la
coopération entre les deux catégories de L permet la production massive d’Ac spécifiques des
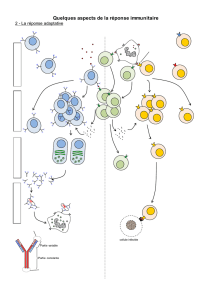
pneumocoques. Ce sont les LT4 spécifiques des pneumocoques qui sont d’abord sélectionnés par les
cellules phagocytaires puis clonés et enfin différenciés en LT4 auxiliaires qui, à leur tour, sélectionnent les
LB spécifiques des pneumocoques. Grâce aux interleukines de ces LT4a le LB spécifiques se clonent puis
se différencient en plasmocytes sécréteurs des Ac anti-pneumocoques.
Corrigé sur les mécanismes de rejet des cellules greffées

II) a) Les deux premières expériences montrent que quel que soit l’âge de la souris nude, son absence de
thymus fait qu’elle accepte la greffe de n’importe quel organe : peau et thymus. Mais une fois qu’elle a
retrouvé un thymus, la greffe de peau est rejetée en 10 jours. Preuve que le thymus est un organe
producteur des cellules effectrices de la lyse du greffon : les LT8 sous leur forme cytolytique.
b) Les cellules du greffon n’ont pas la même identité biologique que le receveur : elles sont repérées par
les cellules sentinelles qui, après les avoir phagocytées, vont recruter les LT4 spécifiques qui se clonent et
se différencient en LT4a, à leur tour recruteurs des LT8 spécifiques qui se clonent aussi et se différencient
en LT cytolytiques (Ltc) tueurs du greffon par le baiser de la mort.
Ainsi si les anticorps (molécules) sont produits par les plasmocytes (issus de la différenciation des
LB) en réponse à la présence d'un antigène et sont spécifiques de ce dernier, le rejet de greffe
repose sur l'action des lymphocytes cytotoxiques (issus des LT8) également spécifiques de
l'antigène (= non soi). Les anticorps contribuent donc à la réaction immunitaire adaptative (=
acquise) à médiation humorale tandis que les LT8 participent à la réaction immunitaire adaptative à
médiation cellulaire
1
/
3
100%