Résumé Rainer

1
Résumé Rainer
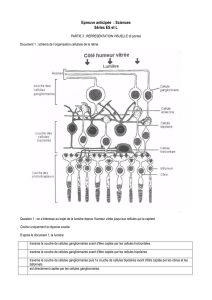
L’épithèle pigmenté fait la première couche de la rétine, cette dernière
absorbe la lumière afin d’éviter des réflexions dans l’œil. Les
photorécepteurs sont de deux types différents, les cônes et les bâtonnets.
Ces dernières vont transformer l’information lumineuse reçu aux cellules
bipolaires qui transmettront aux cellules ganglionnaires, c’est la
transmission verticale. Il y a convergence de l’information, il y a en effet
120mio de bâtonnets et 6mio de cônes pour seulement 1mio de cellules
ganglionnaires.
Entre les couches plexiformes interne et externe, au même niveau que les
cellules bipolaires, se trouvent les cellules amacrine et horizontale qui se
chargent de la transmission horizontale.
La réception de l’information lumineuse se
fait au niveau des cônes et des bâtonnets, ces
dernières contiennent des disques dans lesquels
se trouve la Rhodopsine, qui consiste en une
molécule d’opsine combinée à du rétinal. Le
rétinal combiné à l’opsine est sous forme 11-cis-
Retinal mais va changer de conformation en
absobant un photon devenant all-trans-Retinal
qui se sépare de l’opsine. Elle redeviendra grâce à
une enzyme 11-cis-Retinal, se joindra à nouveau à
une opsine pour être à nouveau prêt.
Dans les cônes il y a trois types de rhodopsine permettant la vision de
couleurs différentes, ces dernières se trouvent dans les plis de la membrane
et non pas dans des disques contrairement aux bâtonnets. Il y a donc trois
types de cônes (vert, rouge et bleu) ayant des spectres d’absorption
différents. Les bâtonnets ont un maximum d’absorption à 500nm de
longueur d’onde, ils servent à percevoir la lumière et non la couleur.
Attention, les couleurs ne sont qu’une interprétation de notre cerveau, la
lumière n’a pas de couleur.
En réalité, le changement de conformation de la rhodopsine va activer la
transducine qui va elle-même activer une phosphodiesterase qui va détruire
le GMPcyclique qui tient ouvert un canal à Na+.
Dans le noir complet, il y a un courant continu entre le segment externe de
la cellule qui fait entrer du Na+ et le segment interne qui fait sortir du K+, la
cellule a un potentiel de -40mV.
Cette réaction en chaine va provoquer une hyperpolarisation de la cellule
car le K+ est toujours pomper dehors. La libération de glutamate
(neurotransmetteur excitateur) est alors inhibée. De plus, il y a amplification
de la réponse à chaque étape, l’activation d’une rhodopsine va activer
plusieurs centaines de transducine qui à leur tour activeront chacune
plusieurs phosphodiesterase, etc.
Les bâtonnets sont 100x plus sensibles que les cônes à la lumière, ils sont
capables de détecter un photon isolé !

2
Les cellules bipolaires sont de deux types : ON et OFF. Elles sont
reliées aux cônes dont elles reçoivent l’information. Les cellules OFF
seront activées par l’absence de lumière de la même façon que les
cônes alors que les cellules ON sont activées par la lumière. Ceci est
possible car elles n’utilisent pas les mêmes types de récepteurs au
glutamate.
Le système horizontale aide dans la vision des contrastes. Ces cellules
vont transmettre des informations de la périphérie.
Exemple : Si le centre est sombre mais les périphéries lumineuses, le
cône central sera actif et les autres autours inactifs, les cellules
horizontales provenant de la périphérie sera alors peu active. Or, ces
dernières libèrent du GABA (neurotransmetteur inhibiteur) lors
qu’elles sont actives. Donc dans ce cas, la cellule centrale ne sera pas
inhibée est sera fortement active alors que si la périphérie était
également sombre, la cellule centrale aurait été inhibée et donc
moins active. Les contrastes ressortent mieux.
Les cellules bipolaires transmettent l’information aux cellules
ganglionnaires fonctionnant également sur un principe ON/OFF qui
emmèneront l’information jusqu’au Thalamus.
Attention, les photorécepteurs, cellules bipolaire et horizontales ne font pas de potentiels d’action, leur réponse est
graduée contrairement aux cellules ganglionnaires qui fonctionne (à partir du premier nœud de Ranvier) avec des
potentiels d’action.
On parle de champ récepteur d’une cellule ganglionnaire pour
parler de la zone de « vision » d’une cellule ganglionnaire. Ces
dernières sont de deux types comme énoncé plus tôt, les cellules
ganglionnaires ON et les OFF. Cependant elles ne reçoivent pas leurs
informations uniquement de cellules bipolaires ON respectivement
OFF. En réalité, les ON ont le centre de leur champ récepteur ON et
les périphérie OFF et réciproquement pour les cellules
ganglionnaires OFF.
Lorsque vous illuminé le centre d’un champ récepteur ON : le centre
est stimulé car sensible à la lumière et la périphérie également car
sensible à l’obscurité. A contrario si on illumine seulement la
périphérie, toutes les zones sont inhibées.
Remarquez cependant qu’à la disparition du stimulus lumineux dans
ce dernier cas, il y a une augmentation de l’activité. La cellule est
sensible aux contrastes (passage de très lumineux à obscure rend ce
dernier encore plus sombre qu’avant) et ceci grâce au système
horizontal.
Jusque-là nous avons parlé des cônes et laissé de côté les bâtonnets.
Ces derniers fonctionnent en effet différemment puisque :
1. Ils ne fonctionnent qu’avec des cellules bipolaires ON et sont
des centaines à donner leur information à une seule cellule
bipolaire.
2. Les cellules bipolaires ON des bâtonnets sont reliées, non
pas aux cellules ganglionnaires, mais à des cellules
amacrines A2

3
3. Ces dernières relient les bâtonnets aux cellules bipolaires ON (par des
gap jonction) ou OFF (par des récepteurs à glycine -> inhibition) des cônes.
L’information est ensuite transmise aux cellules ganglionnaires et plus loin.
Les bâtonnets et les cônes ne sont pas répartis uniformément sur la rétine : La
fovea centralis ne contient que des cônes avec une extrêmement faible
convergence de l’information (1 :1). La foveola, zone la plus interne ne contient
que des cônes pour le vert et le rouge. La macula lutea (tâche jaune) est une
zone pigmentée dû à la photooxydation à cause du manque de vaisseaux
sanguins. Plus l’on va vers la périphérie, moins il y a de cônes par rapport aux
bâtonnets. Dans la périphérie la convergence est très forte. Dans le point
d’entrée du nerf optique, il n’y a pas de photorécepteurs (tâche aveugle).
Il existe deux types de cellules ganglionnaires différentes : les parvocellulaires et les
magnocellulaires. Les parvo servent à la reconnaissance de la forme et de la couleur,
elles ont un petit champ dendritique pour une haute résolution. Elles ne sont pas
capables de sentir les mouvements. Leurs potentiels d’action sont toniques, c-à-d.
qu’il dure dans le temps. A contrario, les magno détectent les mouvements, leur
champ réceptif est grand mais achromatique, elles ont peu de résolution. Leurs
potentiels d’action est transitoire.
On remarque sur la figure à droite que le champ récepteur de la vision des couleurs
fonctionne sur le même principe de contraste qu’expliquer auparavant. Le système
parvocellulaire à donc, en plus de la vision avec centre clair/obscur et périphérie
obscur/clair, une vision avec centre rouge/vert et périphérie vert/rouge ou centre
jaune/bleu et périphérie bleu/jaune. A noter que le jaune est un mélange des
récepteurs rouge et vert, nous n’avons pas de récepteurs préférentiels pour le jaune.
Il faut savoir que tout le processus de vision des couleurs est
travaillé par la suite par notre cerveau qui va nottament prendre
en compte l’éclairage pour ajuster la couleur.
Un autre composant essentiel de la vision est l’adaptation. Ce
mécanisme nous permet de voir tout aussi bien avec une forte
luminance qu’une faible, à une grande distance ou proche. Dans le
noir, nous avons une vision scotopique à l’aide uniquement des
bâtonnets, à la lumière notre vision devient photopique
l’information provient des cônes. La zone entre-deux est dite
mésoscopique, les deux types de récepteurs sont actifs.
Le réflexe pupillaire est le processus
le plus rapide mais également le plus
faible. Il se déroule comme suit :
ganglions cellulaires de la rétine
Prétectum ncl. Edinger Westphal
ganglion cilliaire muscle
sphincter pupillae. Les deux pupilles
sont touchées.
Un mécanisme bien plus efficace
mais prenant plus de temps et celui
d’accumulation dans les récepteurs
(ou vidage) de rhodopsine et

4
transducine. Une cellule exposée longtemps à la lumière deviendra moins
sensible car les molécules seront consommées. À l’inverse, une cellule dans
le noir accumulera la matière et deviendra bien plus sensible. Il faut environ
20min à ce système pour être au maximum. Le point de Kohlraush-Knick
correspond à la sensibilité maximale des cônes.
Le phénomène d’adaptation arrive aussi au niveau local, si on fixe un point
avec une tache lumineuse à droite et une tache sombre au gauche sur un
fond neutre (gris). Les deux taches vont disparaitre progressivement. Si on
reporte notre regard ensuite vers une zone neutre, le « fantôme » des tâches
apparaissent mais dans leur inverse. La cause est que la partie gauche de
l’œil, habituée à l’obscurité verra un contraste
par rapport à avant, et l’inverse pour la partie
droite.
Si l’on suit les voies optiques, les fibres du N.
opticus provenant des hémirétines temporales
restent de leur côté tandis que les fibres
provenant des hémirétines nasales croisent
dans le chiasma optica. Ce croisement permet
d’avoir l’information d’un même endroit par les
deux yeux. Les deux moitiés provenant de deux
yeux après croisement continuent dans le
tractus opticus. Ils arriveront alors au corpus
geniculatum latéral. Ce dernier possède 6
couches distinctes utiles à la rétinotopie,
l’organisation de l’information visuelle. Trois de
ces régions vont prendre l’information
controlatérale et les trois autres l’information
de l’œil ipsilatérale. De plus, l’information des
ganglions magnocellulaires ira aux couches 1-2
de CGL et parvocellulaires aux couches 3 à 6. L’information continue (après cette
séparation) vers le cortex visuel primaire (V1) via les radiatio optica. Parvocellulaires iront
à la couche 4Cα et magnocellulaires à la couche 4Cβ. Il peut survenir des lésions à
n’importe quel endroit sur la voie optique causant des problèmes visuels différents.
On remarque sur les figures à droites un des principes de la rétinotopie. On remarque que
ce sont les plus grandes aires qui travaillent pour les plus petites zones du champ visuel d’un
œil. Mais comme dit auparavant, ce sont les zones où la convergence de l’information est la
plus faible, en d’autres termes l’information provenant du centre est très précise et
beaucoup travaillée.
Les deux hémisphères du cerveau sont en communication par le corpus callosum. La
rétinotopie nous permet de savoir est où interprété l’information du champ
visuel d’une cellule ganglionnaire de la rétine mais également de savoir de
quelle vision un neurone du cortex visuel primaire s’occupe.
Les neurones du cortex visuel primaire sont sélectifs pour l’orientation des
stimuli qui se présentent dans son champ visuel. Ceci se produit par une
intégration qui a lieu dans le CGL.

5
Il faut différencier les cellules simples et complexes dans V1, les
cellules simples possèdent un champ visuel ovale avec une
polarité centrale opposée à ce qui l’entoure (un peu comme les
ganglions ON/OFF). Ce champ récepteur est le fruit de la somme
de ses cellules du CGL au champ récepteur rond.
Figure à droite : 1 : lumière sur ON -> activité. 2 : lumière sur OFF
-> inhibition. 3. Lumière orientée, ON et OFF activés, aucun
changement.
Les cellules complexes sont également sensibles à l’orientation,
ils n’ont cependant pas d’entourage différent est leur stimulation
est tonique contrairement aux cellules simples.
Un autre principe d’organisation régit
V1 : les colonnes d’orientation. Les
neurones des 6 couches du cortex ont
la même préférence d’orientation
suivant une spirale. Si l’on regarde la
spirale depuis le haut, on remarque
que toutes les orientations sont
présentes mais uniquement pour une
partie du champ visuel. La
représentation Pinwheel (comme ici
à droite) fait correspondre une
couleur à une orientation. Sur le
cerveau en entier, on retrouve les
orientations organisées pour le
champ visuel entier.
De plus, si l’on colore avec un marqueur cytochrome-oxydase, on va mettre
en évidence des Blops. Ces derniers contiennent des neurones qui sont
sélectifs pour la couleur et se trouvent aux couches II et III. Les neurones de
ces régions vont travailler l’information en provenance des cellules
ganglionnaires parvocellulaires.
Il existe également des voies de visions extrageniculaires (ne
passant pas par CGL). Le réflexe retino-prétectal de la
contraction de la pupille en est un exemple.
Le système retino-tectal passe par le colliculus superior (CS) et
le pulvinar jusqu’au cortex visuel extrastriaire. Le CS contient
une représentation du champ visuel contralatéral. Ceci
explique la vision aveugle : En cas de lésion totale du CGL (ou
de V1) la personne voit sans s’en rendre compte via ce système.
Il participe également aux commandes de mouvements des
yeux.
Une autre voie, par le noyau suprachiasmaticus, est sensible à
la lumière non pas par les photorécepteurs mais par des
cellules ganglionnaires rétinales spéciales sensible à la lumière.
Cette voie sert au rythme circadien.
Sur la figure de la page suivante, vous avez l’activité de la
couche 4 de V1 de l’hémisphère droit d’un patient après un an
 6
7
8
6
7
8
1
/
8
100%