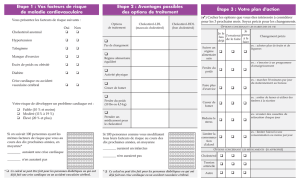
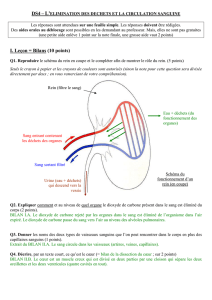
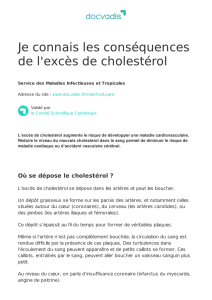
– UE VII : – Biochimie, métabolisme des... 2015-2016 Métabolisme des lipides

2015-2016 Métabolisme des lipides
Nom du cours
– UE VII : – Biochimie, métabolisme des lipides
suite de cours
Semaine : n°8 (du 26/10/15 au
30/10/15)
Date : 29/10/2015
Heure : de 8h00 à
9h00 Professeur : Pr. Brousseau
Binôme : n°45 Correcteur :n°b44
Remarques du professeur
- « je ne suis pas un grand fan de la synthèse des glycérophospholipides et des sphingolipides, sous-
entendu ce n'est pas la peine de trop s'y attarder »
-«la synthèse du cholestérol est beaucoup beaucoup beaucoup plus intéressante »
PLAN DU COURS
II) Métabolisme des AG
A)B) Biosynthèse des AG
B)
3) Synthèse des triglycérides et des glycérophospholipides
- Etape commune
- Synthèse des TG
- Synthèse des Glycérophospholipides ;
a) Synthèse des phosphatidylcholines et des phosphatidyléthanolamines
b) Synthèse des phosphatidylsérines
c) Synthèse des phosphatidylinositols
4) Synthèse des sphingolipides
- Etape commune (synthèse des céramides)
5) Métabolisme du cholestérol
- Biosynthèse du Cholestérol
- Catabolisme du Cholestérol (synthèse des acides biliaires)
1/11

2015-2016 Métabolisme des lipides
II) Métabolisme des AG
C)B) Biosynthèse des AG
3) Synthèse des triglycérides et des glycérophospholipides
1-
1- ÉTAPE
ÉTAPE
COMMUNE
COMMUNE
On regroupe ces deux types de lipides car ils sont construits autour d'un squelette identique : le
glycérol.
Ils vont alors avoir une partie commune de leurs synthèses.
L'un des précurseurs de cette synthèse, pour la partie du squelette (c'est à dire le glycérol) est fourni
pour l'essentiel par la glycolyse, par l'intermédiaire de la 3-P-dihydroxy-acétone.
La 3-P-dihydroxy-acétone va être réduite au niveau de sa fonction cétone en une fonction alcool, (les
électrons étant fournis par NADH2), par la glycérol-3-P déshydrogénase
Cela donne le 3-P-glycérol.
On peut se procurer également du 3-P-glycérol au niveau hépatique, en utilisant le glycérol provenant
du tissu adipeux. En effet au moment de la lipolyse le tissu adipeux libère du glycérol car on mobilise les
acides gras du tissu adipeux, on hydrolyse les liaisons esters entre la partie glycérol et les acides gras du
tissu adipeux. Ce glycérol libéré ne peut alors être estérifié directement pour reformer des triglycérides, il
faut le faire repasser par l'hépatocyte. Une fois dans l'hépatocyte le glycérol va être phosphorylé par un
glycérol kinase pour donner du 3-P-glycérol.
On possède donc deux sources de 3-P-glycérol, soit au niveau de la glycolyse avec la réduction de la 3-
P-dihydroxy-acétone, soit au niveau hépatique en phosphorylant du glycérol.
Les deux fonctions -OH du 3-P-glycérol vont être estérifiées par 2 acides gras, activés par leurs liaison
au CoA, donc par 2 acides CoA, par la glycérol phosphate acyltransférase.
On obtient des molécules de glycérol estérifiées par 2 molécules d'acide gras (qui peuvent être de
nature différente): les Acides Phosphatidiques. Ils peuvent servir comme tel, ou alors comme précurseur
de la synthèse des glycérophospholipides ou des triglycérides.
2/11

2015-2016 Métabolisme des lipides
2- S
2- SYNTHÈSE
YNTHÈSE
DES
DES
TRIGLYCÉRIDES
TRIGLYCÉRIDES
On voit bien que ce qui nous manque, c'est un acide gras sur le carbone 3.Il va donc falloir dans un
premier temps dé-phosphoryler les acides phosphatidiques.
Les acides phosphatidiques peuvent être dé phosphorylés au niveau du carbone 3 par une phosphatase
pour former des Diglycérides.
La fonction OH est remplacée par un acide gras (activé par sa liaison au CoA), grâce à une Diglycéride
acyltransférase qui estérifie la 3 ème liaison permettant d'obtenir des Triglycérides.
3- S
3- SYNTHÈSE
YNTHÈSE
DES
DES
GLYCÉROPHOSPHOLIPIDES
GLYCÉROPHOSPHOLIPIDES
Les glycérophospholipides ont le même squelette que les triglycérides, c'est le glycérol. On part donc
de la fin de l'étape commune de synthèse c'est à dire des acides phosphatidiques.
a) Synthèse des phosphatidylcholines et des phosphatidyléthanolamines
Ici, comme pour les triglycérides, une phosphatase va agir sur les Acides Phosphatidiques, ce qui libère
la dernière fonction OH donnant des Diglycérides. On va alors, non pas comme précédemment fixer un
troisième acide gras mais de la choline ou de l'éthanolamine permettant d'obtenir :
•phosphatidylcholine
•phosphatidyléthanolamine
La liaison de la choline ou de l'éthanolamine avec la CDP leur permet d'être activées et réactives.
L’éthanolamine et la choline arrivent donc sous forme active grâce à leur liaison avec CDP. On va
transférer la partie phospho-choline ou phospho-éthanolamine. Cette réaction libère du CMP.
b) Synthèse des phosphatidylsérines
Les phosphatidyléthanolamines servent de précurseur immédiat à la formation des
phosphatidylsérines grâce à une enzyme échangeuse de base afin d'échanger l'éthanolamine contre de
la sérine.Cette réaction est réversible, on peut donc obtenir de la phosphatidyléthanolamine à partir de
phosphatidylsérine grâce à la même enzyme.
3/11

2015-2016 Métabolisme des lipides
c) Synthèse des phosphatidylinositols
Jusqu'à présent on partait d'un acide phosphatidique et on apporter la choline ou l'éthanolamine
sous forme activée, ici on va faire l'inverse: on va d'abord activer l'acide phosphatidique. Cette activation
se fait grâce au CTP, de celui-ci, se fixe la partie CMP sur le groupement phosphate, ce qui engendre la
formation de CDP-diacylglycérol. On a donc deux groupements phosphate avec de l'énergie au milieu.
Et c'est sur ce CDP-diacylglycérol qui correspond à un diglycéride activé que l'on va pouvoir fixer
l'inositol pour former le phosphatidylinositol.
4) Synthèse des sphingolipides
1-
1- ÉTAPE
ÉTAPE
COMMUNE
COMMUNE (S
(SYNTHÈSE
YNTHÈSE
DES
DES
CÉRAMIDES
CÉRAMIDES)
)
Chez les sphingolipides le squelette n'est plus le glycérol mais la sphingosine.
Pour arriver à la formation de ce squelette on part de molécules très simples:
•1 AA: la sérine
•1 AG à 16C: le Palmityl activé par sa liaison au CoA
Dans une première étape on va condenser la Sérine et le Palmityl CoA. Cette condensation se fait
entre le carbone ( -CO ) de l'acide palmitique et le carbone 2 de la sérine. On libère ici une fonction
carboxylique, donc cette condensation s'accompagne d'une décarboxylation.
Cela ressemble beaucoup dans l'organisation à ce que serait un monoglycéride, c'est à dire à une
molécule qui serait architecturée autour du glycérol et qui porterait une molécule d'acide gras, une
fonction -OH , et une fonction amine. Ce qu'on entend par là, c'est que les glycérophospholipides et les
sphingolipides ont l'air d'être extrêmement différents et c'est le cas d'un point de vue chimique mais en
revanche dans leur organisation dans l'espace ils sont assez proches.
Il y a ensuite une réduction de la fonction alcool puis une déshydrogénation qui forme la double
liaison de la sphingosine. Enfin grâce à une réaction d'amidification on fixe une molécule d'acide gras
sur notre sphingosine. Encore une fois ici, l'acide gras arrive sous forme activée grâce à sa liaison au
CoA. La réaction est assurée par une 4-sphingénine acyltransférase. On obtient un céramide. Les
céramides sont les premiers représentants des sphingolipides.
4/11

2015-2016 Métabolisme des lipides
2-
2- SYNTHÈSE
SYNTHÈSE
DES
DES
DIFFÉRENTS
DIFFÉRENTS
SPHINGOLIPIDES
SPHINGOLIPIDES
On part de la céramide et on occupe la dernière fonction -OH en fixant:
–de la phosphocholine ou de la phosphoéthanolamine pour arriver aux sphingomyélines. La
phosphocholine et la phosphoéthanolamine, on va aller les chercher dans les glycérophospsolipides
correspondant, c'est à dire au niveau des phosphatidylcholines et des phosphatidyléthanolamines.
–du glucose ou ose complexe pour arriver aux gangliosides.
–du glucose ou galactose pour arriver aux cérébrosides puis aux sulfamides. Cette fois ci, les oses sont
activés par une liaison à l'UDP.
5/11
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%