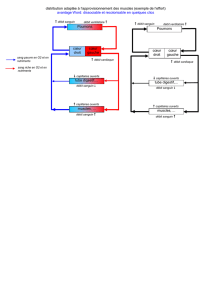
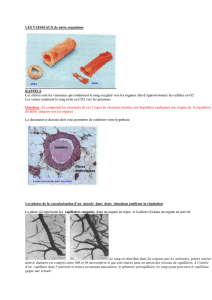
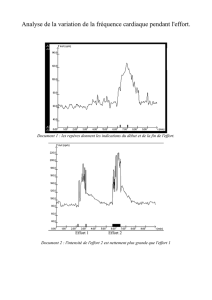
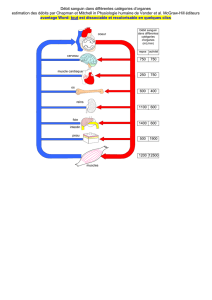
zoologie - E

B22 - Biologie et physiologie animale L1 Bio 2010/2011
- 1 -
ZOOLOGIE
I. Invertébrés
1. Espèce, Eta. Eucaryote et évolution
2. Protistes : protozoaires (rhizopodes, actinopodes, zooflagellés)
3. Protistes : protozoaires (sporozoaires, Microsporidies, infusoires Cilliés)
4. Métazoaires diploblastiques (spongiaires, cnidaires et cténaires)
5. Métazoaires triploblastiques acœlomates (Plathelminthes et Némathelminthes)
6. Métazoaires coelomates Anellides et Mollusques
7. Métazoaires coelomates Arthropoïdes (Arachnides, crustacés)
8. Métazoaires coelomates Arthropodes (Myriapodes et Insectes)
9. Larves d’insectes Echinodermes, Procordés (Amphioxus) Agnathes (Lamproie)
II. Vertébrés
1. Poissons
2. Amphibiens (ou Batraciens)
3. Reptiles
4. Oiseaux
5. Mammifères
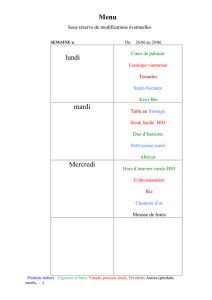
Partie I : L’évolution
A. La classification biologique
Définition : Avant toute étude des êtres vivants, il est nécessaire de pouvoir les CLASSER.
La classification ou la systématique ou la systémique reposent sur les lois dont l’étude porte le
nom de TAXONOMIE.
Le système de la classification biologique date dans sa forme moderne de l’œuvre de
Charles Linné « Systema Natura » de 1735.
→ Système de référence des biologistes.
→ Système synthétique d’une utilisation pratique.
→ Pourrait-on s’en sortir sans un tel système ?
Répartition hiérarchique : Règne, embranchement (Phyllum), classe, ordre, famille, genre et
enfin espèce. Eventuellement on peut parler de race, de variété et d’individu. On peut aussi
rajouter des préfixes tels super- ou sous- (exemple : sous-embranchements).
Qu’est ce qu’une espèce ?
Ce sont des individus qui sont capables de se reproduire entre eux et dont la
descendance est fertile.
Elle est désignée par deux noms latins, le premier correspond au genre et le second
correspond à l’espèce proprement dite. Dans ce cas là la première lettre s’écrit avec une
majuscule et pour le second nom la première lettre commence par une minuscule. Il doit être
en italique ou souligné. Exemple : pour le lion on l’écrit Panthera leo, Oken 1816, FELIN.
Suit le nom de l’auteur qui l’a nommé pour la première fois, l’année de la nomination et le
nom de la famille auquel il appartient.
En bref on a différent systèmes de règnes. Le système mondial accepté par défaut c’est
le système de Whittaker (1969).

B22 - Biologie et physiologie animale L1 Bio 2010/2011
- 2 -
C’est un système avec cinq règnes :
1. Monères (procaryotes)
2. Protistes (eucaryotes unicellulaires)
3. Végétaux
4. Eumycètes
5. Animaux
Ce n’est pas la meilleure classification car n’apparait pas Virus, Prions, etc.
Deux sous-règnes :
1. Protozoaires : animaux unicellulaires
2. Métazoaires : animaux pluricellulaires
Un des critères de bases dans l’étude de l’évolution c’est la notion de triploblastie ou
triploblastique.
Œuf cellule 2 n chromosomes. En bref cet œuf peut se diviser, se segmenter, etc.
Au bout d’un certain temps l’embryon passe différents stades.
En fonction migration cellule on va avoir GASTULATION, alors on peut identifier deux type
de cellule :
-
-
Si elle continue d’évoluer : on a alors présence d’un organisme composé de deux feuillets
Feuillet interne : endodermes
Feuillet externe : ectodermes
On a au milieu une substance gélatineuse
L’ouverture s’appelle Blastopore
Diploblastie = éponge, méduse, etc.
Si continue alors apparition d’un troisième feuillet embryonnaire. Le MESODERME
Trois feuillet alors TRIPLOBASTIE
Dernière étape, si ce mésoderme reste tel quel, c’est à dire qu’il ne va pas s’ouvrir, ne va pas
produire de CEDOME ; Alors il est appelé ACCELOMATES
Et si ce mésoderme se sépare à nouveau et donne une caractéristique alors c’est
CEOLOMATES
Deux feuillets = DIPLOBLASTIE
Trois feuillet = TRIPLOBASTIE
B. L’évolution des premières cellules
Passage de l’état procaryote à l’état eucaryote.
Origine de la vie :
Il n’y a pas de consensus général pour une théorie de l’origine de la vie
Malgré cela il existe des caractéristiques communes en particulier si on considère qu’elle peut
provenir d’autres planète.
Donnée fondamentale : origine vers 3.8 3.5 Milliard d’année
Théorie classique d’Oparie Haldane : production rapide de molécules sur la terre suivie de
réaction chimique qui on produit une complexité organique croissante jusqu’à obtenir une vie
organique capable de se reproduire de muter et de subir la sélection en matière organique
comme nutriment.

B22 - Biologie et physiologie animale L1 Bio 2010/2011
- 3 -
Autre théorie : la vie vient d’ailleurs par des processus d’impact de planète (Arrhenius 1908
par des spores extraterrestres) spores ou bactéries par météorites voyages dans l’espace (a été
démontré) ou par d’autre être intelligent volontairement. Seulement en présence d’eau.
Autre théorie : sur terre à partir de matière inorganique.
Chronologie des premiers instants après le Bing Bang (ce ne sera pas à l’interrogation) :
1. Zéro seconde : Bing Bang
Explosion primordiale : elle est accessible aux maths (topologie, elle étudie les propriétés
géométriques invariantes d’un objet quand celui-ci est étiré, tordu ou rétrécie de manière
continue)
Cette explosion primordiale est inaccessible pour la physique !
Nouvel âge de l’univers : il y a 16 à 17 milliards d’années. Naissance présumé de l’univers,
puisque la lumière la plus ancienne qui nous parvient a parcouru une distance de 17milliard
d’année lumières (selon les modèles cosmologique l’âge de l’univers varie de 9 à 20milliard
d’année.
C’est l’ère de la super force (hypothèse) l’univers est sous l’influence de la super gravité
(gravité quantique).
Elle unifie les 4 forces fondamentales qui apparaitront plus tard : nucléaire forte, nucléaire
faible, électromagnétique et gravitationnelle (ces 4 sont démontrées). Chaque force est
associée à une famille de particules.
A ce stade d’évolution, la matière n’était pas encore née, seul le « vide » régnait.
Le vide de l’univers n’était pas vide !!
2. 10^-43 seconde après le Bing Bang : période dite de grande unification.
La super force se scinde en deux forces (hypothèse car se sont les deux forces qu’on a réussi à
reconstituer) la gravitation et le force électronucléaire.
La force électronucléaire regroupe les interactions fortes et électrofaibles. Elle est décrite par
la théorie actuelle de la grande unification ou TGU.
10^-43 sec correspond à ce que l’on dénomme le temps de Planck unité de temps
incompressible.
Cet intervalle de temps semble être le plus petit possible selon la physique quantique de la
même manière que la distance de Planck (10-35 m) semble être la plus petite distance
accessible à notre physique.
Le cosmos est donc d’environs 10-35m.
3. 10^-35 sec inflation et séparation des forces.
7. 10^-6 sec à une seconde
La matière est visible et les maths laissent place à la physique.
9. 3 minutes
Baisse de l’énergie de la température qui se poursuit lentement, matière se stabilise.
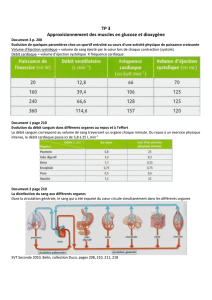
L’évolution (à savoir) :

B22 - Biologie et physiologie animale L1 Bio 2010/2011
- 4 -
Si on considère 4,5milliard d’année, au niveau de l’environnement et de l’oxygène, on
s’aperçoit que la période charnière se situe autours de 2milliard d’année, avant on avait un
environnement sans oxygène, ce n’était pas stable, en revanche à partir de 2milliard d’année
on a une situation riche en oxygène et surtout stable. Les procaryotes, lorsque l’oxygène
n’étaient pas stables existaient déjà vers 3,5milliard d’année, cependant à partir de 2milliard
d’année, se sont développés les premiers eucaryotes à partir des algues.
C’est ce passage clé qui fait qu’on passe des procaryotes aux eucaryotes. Il n’y a plus
vraiment d’évolution possible pour les procaryotes contrairement aux eucaryotes ce qui
explique beaucoup de chose concernant l’évolution.
Organismes pluricellulaires :
Quaternaires : évolution de l’homme
Cénozoïque : tertiaire : radiation des mammifères
Mésozoïque :
Crétacé 100millions d’années: derniers dinosauriens premiers primates premières plantes à
fleurs (angiospermes) 65millions d’années fin du crétacés.
Jurassique 160millions d’années : premiers oiseaux et dinosauriens
Trias 200millions d’années : premiers mammifères
Paléozoïque :
Westphalien 300millions d’années : premiers reptiles
Dévonien 360millions d’années : premiers amphibiens
Cambrien 550millions d’années : premiers poissons
Précambrien
(chercher schéma sur internet)
Hypothèse de la disparition des dinosaures :
1. Trop nombreux et trop gros
2. Climat
Volcans émission de souffre acidification de l’atmosphère donc modification des végétaux,
moins de nourriture, etc.
Ce que l’on sait c’est que les dinosaures ont disparu lentement.
3. Evolution
Changement physique de la planète, les océans montent et descendent avec le réchauffement
et la tectonique des plaques d’environs + ou Ŕ 200m
4. Astéroïde
Ca a été brutal donc on pense que ce n’est pas la cause principale de la disparition des
dinosaures mais elle aurait donné le coup final de la disparition des dinosaures.
Ce processus de disparition était inéluctable. Tout a été bouleversé suite à leur disparition.
A la fin du crétacé c’est ce qu’on appelle une période d’extinction des espèces, chute de la
biodiversité. Les dinosaures disparaissent, les crocodiles il en reste peu, mammifère 75%,
lézard et serpent augmentes, les tortues survivent, les oursins 71% survivent, bivalves 92%
plancton végétale 47%.
Trias 150millions d’années, apparitions des dinosaures, ils se développent jusqu’à 65millions
d’années et ils disparaissent et laissent place à l’évolution des autres espèces. Les mammifères
avant 65millions d’années étaient peu nombreux et tout petit, moins de 10kg, donc tant qu’il y

B22 - Biologie et physiologie animale L1 Bio 2010/2011
- 5 -
avait les dinosaures l’évolution des mammifères étaient impossibles. Lorsque les dinosaures
disparaissent, les mammifères peuvent se développer. Ils se développent dans les niches
écologiques libérées par les dinosaures.
Aujourd’hui, on a identifié 1,750 millions d’espèces (ou 2millions).
Animaux : 1 320 000, Protozoaires : 80 000, Plantes : 0,3 millions, Insectes : 0,9 millions.
Virus, on en connait peu mais on estime à 0,4 millions ceux qui existent et qu’on a toujours
pas identifié, de même pour les bactéries, on estime à 1,1millions les bactéries non identifié.
On estime les insectes à 8millions.
Bilan actuel : on est à 2millions d’espèces connues au lieu de 20millions d’espèces supposées
voir 100millions d’espèces sur la planète avec les fonds océaniques.
Mise en évidence processus évolution :
- Paléontologie et anatomie comparée.
- Informations phylogénétiques dans le génome.
- L’hypothèse actuelle très controversé c’est que tous les organismes vivants ont
une origine commune. Un auteur WOESE et FOX en 1977 ont parlé et proposé
une cellule virtuelle qu’on appelle cellule ancestrale ou le progénote.
Pourquoi cette théorie ? Elle a été proposée tout simplement parce que quand on fait du
séquençage d’ARN ribosomique sur par exemple une souris, du riz, des protistes et des
bactéries, on s’aperçoit qu’une fois le séquençage effectuée, ces organismes complètements
différents possèdent environs 80% à 99% de ressemblances.
Etude des distances statistique entre les différents groupes va permettre la différenciation des
espèces et leur évolution les uns par rapport aux autres. Ainsi on en conclut que les bactéries
par exemple sont plus éloignées que les animaux et les plantes.
C’est la théorie de l’évolution qui est actuellement acceptée.
Théoriciens de l’évolution :
1. Empédocle d’Agrigente (5 av. J.C.) : scientifique avant tout mathématicien. Puis il
s’est lancé dans la biologie. Comment expliquer qu’on en soit là ? Celui-ci dit qu’à un
certain moment la terre s’est mise à produire une multitude d’organes séparés (tête,
jambe, bras, etc.) puis ils se sont assemblés et ont formé les êtres vivants que l’on
connait actuellement (grâce aux Dieux etc.) d’où les différentes espèces.
2. Aristote (4 av. J.C.) : explique qu’il n’en est pas sûr mais il constate qu’il y a une
analogie entre les jambes les nageoires et les ailes, les arrêtes et les os. Il pense qu’une
notion de temps a eu lieu en sous entendant l’évolution mais ne prononce pas le mot. Il
est le premier à donner la notion.
3. Lucrèce (1 av. J.C.) : théorie des atomes et des molécules qui constituent la matière,
c’est un disciple d’Epicure. Pour lui la roches les troncs, etc. sont constitués par des
« choses » beaucoup plus petites. Il dit que tout ce qui existe a toujours été tel quel et
qu’il n’y a pas d’évolution.
4. Buffon (19ème siècle) : un naturaliste bourguignon c’est le premier à évoquer une
origine commune entre tous les animaux. Ce pendant les autres lui disent que si nous
avions une origine commune avec les animaux, pour l’homme et le singe par exemple,
ça ne colle pas du tout.
Buffon : le premier à parler d’un ancêtre commun (19ème siècle)
Deux grands théoriciens de l’évolution : Lamarck et Darwin
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
1
/
59
100%