3 Le brassage génétique par la méiose et la fécondation

Terminale S 19/56
Partie 3 – Stabilité et variabilité des génomes - Évolution Rev. 665 du 06/11/2009 15:38
3 Le brassage génétique par la méiose et la
fécondation
3.1 Correspondance phénotype/génotype
Elle est simple chez les organismes haploïdes : un caractère correspond
au seul allèle présent.
3.1.1 Mise en évidence par test-cross
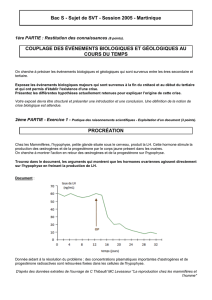
[130-131]À cause de la dominance, pour un gène étudié, d’un allèle par
rapport à d’autres, le génotype ne peut pas être déduit du phénotype chez
les organismes diploïdes.
[132.A1]Le croisement-test (test-cross) consiste à croiser une population
homozygote et récessive pour le caractère à étudier, avec la population à
tester. Les individus de la souche-test sont issus de lignées pures,
obtenues par des croisements répétés.
[131.B4]Le test-cross est inutile dans le cas d’une codominance ou d’une
dominance incomplète.
3.1.2 Analyse d’arbres généalogiques
[133.B2]Le test-cross ne peut être effectué que sur des descendances
importantes. Chez l’homme, l’analyse d’arbres généalogiques permet de
déduire le génotype des membres d’une famille, à condition d’étudier
plusieurs générations.
3.2 Le brassage interchromosomique
3.2.1 Dû à la méiose
[135.B3]Chaque gamète ne contient pas la même information génétique,
suite à la ségrégation indépendante des chromosomes homologues lors de
la première division de méiose. Le nombre de distributions possibles des
chromosomes est 2n.
Le brassage interchromosomique est à l’origine de phénotypes nouveaux.
Le croisement-test sur une génération F1 issue du croisement de 2 lignées
pures (>=2 gènes indépendants) donne : 25% pour chaque phénotype
parental, 25% pour chacun des 2 phénotypes nouveaux.
3.2.2 Dû à la fécondation
La rencontre des gamètes se fait au hasard. Donc pour un couple donné,
les combinaisons possibles de chromosomes chez le zygote sont de 22n.
La variabilité génétique est d’autant plus forte que :
le nombre de gènes sur chaque chromosome est élevé

Terminale S 20/56
Partie 3 – Stabilité et variabilité des génomes - Évolution Rev. 665 du 06/11/2009 15:38
l’hétérozygotie est importante.
3.3 Le brassage intrachromosomique
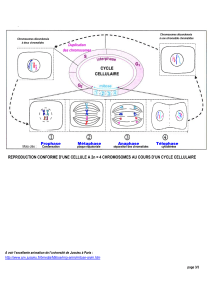
3.3.1 Observation
[137.B3]Pendant la prophase 1, les chromosomes homologues forment des
tétrades. Les chromatides homologues s’enchevêtrent, les points de
croisement étant appelés chiasma. Des ruptures de l’ADN peuvent
intervenir au niveau des chiasmas. Des portions de chromatides sont alors
échangées entre les deux chromosomes (crossing-over ou enjambement).
[139.B3]Les chromosomes issus de ce brassage sont dits recombinés :
chacun des chromosomes homologues ne contient plus l’information qu’il
possédait avant la méiose. Les chromatides ne portent pas la même
information génétique (sur une portion +/- grande)
3.3.2 Conséquences
Dans le cas de gènes liés, des phénotypes non-parentaux apparaissent en
F2 (croisement-test). Ils sont identiques à ceux qui apparaîtraient si les 2
gènes étaient indépendants. [136.A1]La fréquence des différents phénotypes
n’est cependant pas équiprobable (%phénotypes parentaux >
%phénotypes recombinés)
Les crossing-over ne touchent qu’une seule des deux chromatides : le
pourcentage de recombinaison maximum est de 50% (1 chromatide sur 2
totalement recombinée). Les chiasmas peuvent se placer à n’importe quel
niveau.
Le nombre de crossing-over est limité pour chaque tétrade, à cause de la
rigidité des chromosomes. Deux conséquences :
La proportion de gamètes recombinés sera d’autant plus forte que la
distance entre les 2 gènes étudiés sur la chromatide est importante.
Si la distance entre ces deux gènes est suffisamment grande, deux
crossing-over peuvent intervenir simultanément : on retrouve alors,
pour ces deux gènes, des gamètes de type parental.
3.4 Bilan
1
/
2
100%