12 à 24h

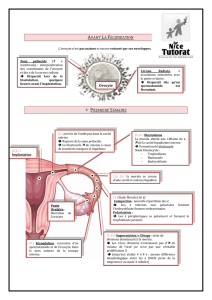
I mplantation


Morula 3,5 jours
Blastocyte 5 jours
Fécondation :
12 à 24h
Avant l'implantation
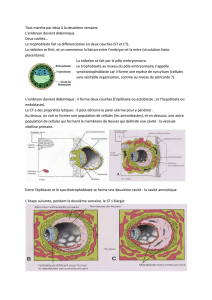
À ce stade, la morula est devenue
blastula ou blastocyste.
À cette fin, les cellules du blastocyste se
différencient, se déplacent et se
réarrangent pour former:
* les tissus embryonnaires (les feuillets
germinaux) ;
* les tissus extraembryonnaires (les structures
de soutien).
Au pôle apical du blastocyste se forme un amas de
cellules internes que l'on appelle l'embryoblaste ou
bouton embryonnaire. C'est là que commence la
genèse de l'embryon.
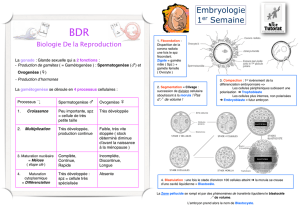
La première division se fait selon un plan
méridional passant par les pôles du
zygote. Ensuite, un des deux blastomères
résultants se divise également selon un
plan méridional. L'autre blastomère, par
contre, se divise selon un plan équatorial.
Au fur et à mesure des divisions
successives les cellules, restant
agglutinées, forment la morula. Les
cellules externes de celle-ci forment le
trophoblaste qui constituera la couche
superficielle du placenta. Les cellules
internes s'agglutinent à un pôle de la
morula et forment la masse cellulaire
interne ainsi qu'une cavité, le blastocèle.

A partir du 8ème jour, la cavité
amniotique apparait dans
l’épiblaste: «lapartie épaisse de
la paroi du germe qui avait
établit le premier contact avec
l’utérus , fait apparaitre trois
zones distinctes: une zone
externe, une zone interne et une
intermédiaire entre les deux.
Dans cette dernière, on observe
une nouvelle exsudation de
liquide. Nous appelons ce liquide
l’eau dorsale du blastème, en le
différentiant de l’eau ventrale du
blastème, apparue
antérieurement» (EB)

Lieux d'Implantation
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
1
/
32
100%