la fecondation - DSI friends 972

!"#$%&'()"*+'(##
,- +./01234/51.##
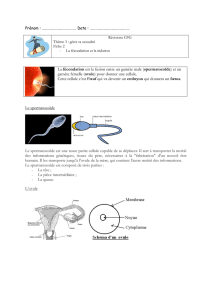
!67#0648604867#09:;57967#467#260.5<067#:..967#1./#=1./09#>36#2?3.#.9=:/126#@#;?81==6A#
;67#B01467737#26#29C6;1BB6=6./#06B176./#730#;67#=D=67#:4/5C5/97#46;;3;:5067#E1.2:=6./:;67F#
!:#41.7/5/3/51.#2?3.#6=G0H1.#A#B357#2?3.#:23;/6#.946775/6#>36#;67#46;;3;67#B01;5E<06./#A#
41==3.5>36./#6./06#6;;67#A#7?5.E10=6./##26#;630#B175/51.#A#48:.I6./#26#E10=6#A#76#29B;:46./#A#
76#06I013B6./#A#76#25EE906.456./#A#6/#2:.7#460/:5.7#4:7#06./06./#6.#:B1B/176#JK#=10/#46;;3;:506#
B01I0:==96-#F##
*13/67#767#:4/5C5/97#71./#411021..967#2:.7#;?67B:46#6/#2:.7#;6#/6=B7#26#=:.5<067#/0<7#
B0945767#6/#71./#7137#;6#41./0L;6#267#I<.67#23#29C6;1BB6=6./F##
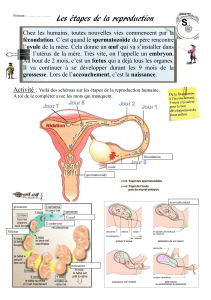
'.#257/5.I36#:3#41307#23#29C6;1BB6=6./#M#9/:B67#N#
O;:#E941.2:/51.##
O#;:#76I=6./:/51.##
O;:#I:7/03;:/51.##
O;:#.6303;:/51.##
O;?10I:.1I9.<76#
&8:43.6#267#9/:B67#67/#=:0>396#B:0#267#9C<.6=6./7#46;;3;:5067#6/#=1;943;:5067#.13C6:3PA#
>35#B63C6./#7?:441=B;50#>36#75#;67#9/:B67#B094926./67#1./#9/97#09:;57967#410064/6=6./F##
!:#E941.2:/51.#C:#=125E560#B01E1.29=6./#;:#7/034/306#26#;?Q3E#6/#41.25/51..6#;6#
29C6;1BB6=6./#E3/30#26#;?6=G0H1.F###
!:#76I=6./:/51.#/0:.7E10=6#;?Q3E#6.#3.6#7/034/306#B;30546;;3;:506F#
!:#I:7/03;:/51.#41./05G36#:3#B175/51..6=6./#267#R#E635;;6/7#6=G0H1..:5067#B05=1025:3PA#6/#
6;;6#C:#D/06#:486C96#:3#41307#26#;:#.6303;:/51.F##
&6//6#9/:B6#E:5/#:3775#;?1GS6/#26#;?:=1046#23#29C6;1BB6=6./#26#;?6=G0H1.#>35#C:#7?:;;1.I60#
76;1.#;?:P6#:./901OB17/905630#6/#7#:B;:/50#76;1.#3.#:P6#21071OC6./0:;F##
+;#H#:30:#:3775#=576#6.#B;:46#2?3.#/3G6#4063P#2:.7#;:#09I51.#2107:;6F##
!#10I:.1I9.<76#41.7/5/36#;?9/:B6#13#;67#10I:.67#6/#;67#/57737#B06..6./#;6307#B;:467#
29E5.5/5C67#6/#C1./#76#25EE906.4560A#B357#411021..6./#;630#E1.4/51..6=6./#B130#41.7/5/360#3.#
10I:.57=6#:3/1.1=6F##
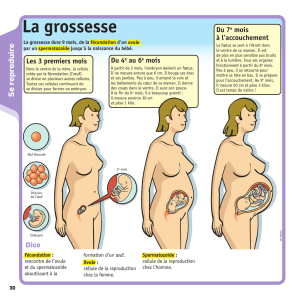
T- U6.41./06#267#I:=</67#

!:#E941.2:/51.A#4?67/#;6#B89.1=<.6#26#E3751.#23#I:=</6#=:;6A#26#B6/5/6#/:5;;6A#/0<7#=1G5;6#
>3?67/#;6#7B60=:/1V1W26A#:C64#3.#I:=</6#E6=6;;6A#>35#;35#:3#41./0:506#67/#/0<7#C1;3=5.63P#6/#
5==1G5;6F###
&6//6#E3751.#21..60:#3.6#46;;3;6#3.5>36A#;?Q3EA#>36#;?1.#:BB6;;6#;6#VHI1/6A#4?67/#;6#B15./#26#
29B:0/#23#29C6;1BB6=6./#2?3.#.13C6;#10I:.57=6F##
!:#E3751.#267#=:/9056;7#I9.9/5>367#67/#25/6#:=B85=5P56A#:3#41307#26#46//6#E941.2:/51.#
;?1C3;6#67/#:;107#:4/5C9#6/#;:#46;;3;6#0626C56./#25B;1W26F#+;#H#:#267#=94:.57=67#26#41./0L;67A#26#
;?6./096#26#7B60=:/1V1W267A#>35#C1./#;5=5/60#;:#E941.2:/51.#6./06#67B<467#25EE906./67F##
!?5./60:4/51.#6./06#;6#7B60=:/1V1W26#6/#;:#730E:46#26#;?1C14H/6#B01C1>36#3.6#4:74:26#26#
09:4/51.7#=6=G0:.:5067A#4H/1B;:7=5>36#6/#.34;9:5067#>35#41.23576./#@#;?:4/5C:/51.#23#
=9/:G1;57=6#26#;?Q3EA#=:57#:3775#:3#G;14:I6#26#;:#B1;H7B60=56#6/#:#;?5.5/5:/51.#23#
B01I0:==6#26#29C6;1BB6=6./F##
!67#09:4/51.7#25/67#2?:4/5C:/51.#=125E56./#6.#B01E1.2630A#;:#7/034/306#4H/1B;:7=5>36#26#
;?Q3EA#;:#257/05G3/51.#26#460/:5.7#"U(#=677:I607A#6/#46;:#6./0:5.6#;:#257B:05/51.#26#;:#7H=9/056#
0:25:;6#26#;?1C14H/6#B130#;:57760#B;:46#@#3.6#7H=9/056#G5;:/90:;6F#
&86V#;67#3012<;67#J/05/1.A#7:;:=:.206-#:3#=1=6./#26#;?:4413B;6=6./#;67#7B60=:/1V1W267#
71./#4:B/67#B:0#;:#E6=6;;6#6/#C1./#D/06#7/14X97#2:.7#;6#4;1:>36F#
!:#E941.2:/51.##>35#C:#7?5.5/560#:3#.5C6:3#23#4;1:>36#C:#76#B01;1.I60#;107#26#;:#B1./6A#1.#
B:0;6#26#E941.2:/51.#B76321#6P/60.6F##
&86V#;67#"('YUZA#;:#E941.2:/51.#76#E:5/#8107#26#;?10I:.57=6#=:/60.6;A#6;;6#67/#25/6#6P/60.6F#
):.7#46#4:7#:3#=1=6./#26#;:#B1./6A#;6#7B60=:/1V1W26#06./06#6.#41./:4/#:C64#3.#41021.#
I9;:/5.63P#>36#;?1.#:BB6;;6#;:#I:.I36A#B357#:C64#;:#41>36A#B357#:C64#;?6.C6;1BB6#C5/6;;5.6F##
&67#5./60:4/51.7#71./#B05=1025:;67#4:0#6;;67#41.E<06./#:3#7B60=:/1V1W26#;:#4:B:45/9#26#
E941.260#;?1C14H/6#TF#&67#5./60:4/51.7#B05=1025:;67#71./#7B945E5>367#26#;?67B<46#6/#;5=5/6#
21.4#;67#E941.2:/51.7#40157967F##
&86V#;67#=:==5E<067A#;67#1G760C:/51.7#=5401741B5>367#5.25>36./#>36#763;7#;6#.1H:3#6/#;6#
46./051;6#B01P5=:;#23#7B60=:/1V1W26#B9.</06#2:.7#;6#4H/1B;:7=6#26#;?1C14H/6F#
'.#:BB6;;6#46./051;6A#;6#46./06#26#.34;9:/51.#:#B:0/50#23>36;A#;67#=5401/3G3;67#B1;H=90576./#
B130#E10=60#3.#:7/60A#>35#2:.7#;6#4:7#23#7B60=:/1V1W26#B10/6#3.#.1=#7B945E5>36#N#;6#
7B60=:7/60F##
!67#=5401/3G3;67#23#7B60=:/1V1W26#C1./#06./060#6.#41./:4/#:C64#;67#=5401E5;:=6./7#23#
410/6P#26#;:#E3/306#09I51.#2107:;6#26#;?Q3EF#&6#7H7/<=6#26#=5401/3G3;67#76=G;6#B01=13C150#
;6#29B;:46=6./#267#T#.1H:3PA#=[;6#6/#E6=6;;6#J;67#B01.34;937-#C607#;:#09I51.#46./0:;6#26#
;?89=57B8<06#:.5=:;#13#5;7#C1./#E3751..60#B130#067/:3060#;:#25B;1W256F##

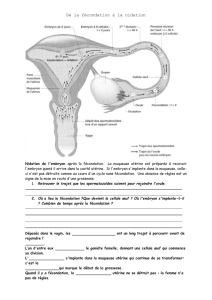
!?5./60:4/51.#6./06#;6#7B60=:/1V1W26#6/#;:#730E:46#26#;?1C14H/6#B01C1>36#3.6#4:74:26#26#
09:4/51.7#:3#.5C6:3#=6=G0:.:506A#4H/1B;:7=5>36#6/#.34;9:506F#*13/#4645#C:#41.23506#@#
;?:4/5C:/51.#23#=9/:G1;57=6#26#;?Q3E#=:57#:3775#;6#G;14:I6#26#;:#B1;H7B60=56#6/#;?5.5/5:/51.#23#
B01I0:==6#26#29C6;1BB6=6./F#/13/67#467#09:4/51.7#71./#:BB6;967#09:4/51.7#2?:4/5C:/51.#F#
R- U910I:.57:/51.#23#4H/1B;:7=6#2:.7#;?Q3E#
"B067#;:#E3751.#6./06#;67#=6=G0:.67#B;:7=5>367#267#7B60=:/1V1W267#6/#267#1C14H/67A#;:#
=6=G0:.6#26#;:#46;;3;6#E941.296#C:#76#29B1;:05760#/0<7#0:B526=6./A#S37/6#:C:./#26#76#
06B1;:05760#>36;>367#5.7/:./7#:B0<7F#+;#7?:I5/#2?3.6#=125E54:/51.#/0<7#0:B526#23#B1/6./56;#26#
=6=G0:.6A#>35#C:#0923506#;:#B1775G5;5/9#26#E3751.#2?3.#.13C6:3#7B60=:/1V1W26#:C64#
;?1C14H/6F##
!:#=125E54:/51.#0:B526#23#B1/6./56;#26#=6=G0:.6#C:#0923506#;67#48:.467#26#E941.2:/51.#
:C64#3.#7641.2#7B60=:/1V1W26A#4?67/#;:#B06=5<06#G:005<06#:#;:#B1;H7B60=56A#4?67/#3.6#
G:005<06#26#/HB6#51.5>36F##
!:#E941.2:/51.#C:#:3775#B01C1>360#267#=125E54:/51.7#26#E;3P#51.5>367#@#/0:C607#;:#=6=G0:.6#
B;:7=5>36A#6/#C:#B01C1>360#3.6#;5G90:/51.#26#4:;453=#5=B10/:./6#2:.7#;6#4H/1B;:7=6#JB:0#;6#
09/543;3=#6.21B;:7=5>36A#6/#;67#=5/1481.20567-#F%;;6#:30:#B130#41.79>36.46A#;:#41./0:4/51.#
23#410/6P#26#;:#46;;3;6A#4:#>35#C:#29B;:460#;67#I0:.3;67#410/54:3PF#
&67#T#=6=G0:.67#:005C6./#6.#41./:4/#6/#E3751..6./A#;67#I0:.3;67#410/54:3P#C1./#21.4#
7?13C050A#6/#;6#41./6.3#67/#;5G909#6./06#;:#=6=G0:.6#26#;:#46;;3;6#6/#;?6.C6;1BB6#C5/6;;5.6F#&6#
B89.1=<.6#76#B01B:I6#/13/#:3#;1.I#26##;:#730E:46#46;;3;:506F#
!#6P14H/176#26#I0:.3;67#410/54:3P#:#263P#41.79>36.467#N#
O5;#H#:30:#E10=:/51.#2?3.#67B:46#6./06#;:#=6=G0:.6#6/#;?6.C6;1BB6#C5/6;;5.6A#46/#67B:46#B10/6#
;6#.1=#2?67B:46#B09C5/6;;5.F#
O5;#H#:30:#=576#6.#B;:46#2?3.6#=6=G0:.6#26#E941.2:/51.F#
\=576#6.#B;:46#26#;?67B:46#B09C5/6;;5.#N#
]:0=5#;67#41.7/5/3:./7#267#I0:.3;67#410/54:3PA#1.#06/013C6#267#=341B1;H7:448:05267#>35#:3#
41./:4/#23#=5;563#6P/905630#C1./#E10=60#3.#I6;#8H201B85;6F#!?8H20:/:/51.#6#467#
=341B1;H7:448:05267#C:#6.I6.2060#3.#:EE;3P#26#;5>3526#6./06#;:#=6=G0:.6#B;:7=5>36#6/#
;?6.C6;1BB6#C5/6;;5.6F#
+;#H#:#:3775#267#6.VH=67#>35#:3#41./:4/#23#=5;563#6P/905630#C1./#D/06#:4/5C967A#6/#C1./#
8H201;H760#;67#5./60:4/51.7#=1;943;:5067#>35#6P57/6./#6./06#;?6.C6;1BB6#C5/6;;5.6#6/#;:#730E:46#
26#;:#46;;3;6F#

"3#E5.:;#;?:4/5C5/9#6.VH=:/5>36#267#6.VH=67#6/#;?:EE;3P#26#;5>35267#C1./#B01C1>360#;6#
713;<C6=6./#26#;?6.C6;1BB6#C5/6;;5.6#B:0#0:BB10/#@#;:#730E:46#26#;:#=6=G0:.6#B;:7=5>36#
B130#B01C1>360#;:#E10=:/51.#26#;?67B:46#B905OC5/6;;5.F#
\=576#6.#B;:46#26#;:#=6=G0:.6#26#E941.2:/51.##
Y.6#260.5<06#E0:4/51.#23#41./6.3#267#I0:.3;67#410/54:3P#67/#E10=6#26#I;H41B01/95.67#>35#
/0:C6076./#;?6.C6;1BB6#C5/6;;5.6A#76#B1;H=90576./A#6/#41.7/5/36./#3.#:=:7#26#I;H41B01/95.67#:#
;:#E:46#6P/60.6#26#;?6.C6;1BB6F#
!:#=6=G0:.6#67/#:5.75#=125E596#6/#6;;6#26C56./#=6=G0:.6#26#E941.2:/51.#>36#;?1.#:BB6;;6#
:3775#;6#481051.F#"3#E5.:;#;:#E10=:/51.#23#481051.#6/#26#;?67B:46#B09C5/6;;5.#41.7/5/36./#3.6#
7641.26#G:005<06#@#;:#B1;H7B60=56A#6/#21.4#=576#6.#B;:46#2?3.6#7641.26#G:005<06#B8H75>36F##
!:#=125E54:/51.#26#;:#7/034/306#26#;?6.C6;1BB6#C5/6;;5.6#:5.75#>36#;:#E10=:/51.#26#;?67B:46#
B09C5/6;;5.#1./#3.6#:3/06#41.79>36.46A#6;;67#C1./#06.206#;?Q3E#;5G06#2:.7#767#6.C6;1BB67A#;6#
4H/1B;:7=6#:30:#:;107#73G5#267#=125E54:/51.7#6.#/60=67#26#=13C6=6./F#
%.#6EE6/A#;?Q3E#C:#7?1056./60#7137#;?6EE6/#26#;:#B67:./630A#7137#;?6EE6/#2?3.#I0:256./#C5/6;;5.A#26#
/6;;6#710/6#>36#;67#Q3E7#E941.297#C1./#/137#B0976./60#;?89=57B8<06#:.5=:;#C607#;6#8:3/F##
&6//6#01/:/51.#2?1056./:/51.#67/#;6#B06=560#75I.6#6P/60.6#>36#;?1C14H/6#T#67/#E941.29F#&6#
B89.1=<.6#67/#1G760C9#6.C501.#R^#=.#:B0<7#;:#E941.2:/51.F#!?6.76=G;6#26#467#09:4/51.7#
41.7/5/36./#;67#09:4/51.7#2#:4/5C:/51.7F#
):.7#;?89=57B8<06#:.5=:;#;:#06B0576#26#;:#=95176#6./0:5.6#@#;?9=57751.#23#7641.2#I;1G3;6#
B1;:506#6/#;:#257B:05/51.#23#E376:3#=5/1/5>36F#+;#H#:30:#:;107#6P/6.751.7#23#B5I=6./#2:.7#;6#
410/6P#:3O267737#23#.1H:3A#21.4#;:#/:486#26#=:/30:/51.#C:#257B:0:5/06F##
&6//6#257B:05/51.#41.7/5/36#;6#263P5<=6#75I.6#6P/60.6#>35#B013C6#>36#;:#E941.2:/51.#67/#
09:;5796F#!:#01/:/51.#23#410/6P#C:#6.I6.2060#;:#=576#6.#B;:46#2?3.#B;:.#26#7H=9/056#6/#2?3.#
.13C6;#:P6F#
_- U1/:/51.#23#410/6P#N#=576#6.#B;:46#26#;:#7H=9/056#6/#2?3.#.13C6;#:P6#
:- !:#01/:/51.#410/54:;6#
#
!6#260.560#9C<.6=6./#67/#;:#01/:/51.#410/54:;6##13#01/:/51.#26#7H=9/057:/51.F#%;;6#:#;563#
6.C501.#3.6#,`R^#:B0<7#;:#E941.2:/51.A#;6#410/6P#73G5/#3.6#01/:/51.#26#R^a#6.#25064/51.#23#
B15./#26#B9.9/0:/51.#23#7B60=:/1V1W26A#:;107#>36#;6#4H/1B;:7=6#B01E1.2#26=6306#
5==1G5;6F#
!?:.:;H76#26#;:#;14:;57:/51.#26#;:#/3G3;5.6#J41.7/5/3:./#=:S630#267#
=5401/3G3;67#K29B;:46=6./#26#;?Q3E-A#=1./06#>3?:C:./#;:#01/:/51.#410/54:;6A#267#E:5746:3P#
26#=5401/3G3;67#76#=6//6./#6.#B;:46A#5;#H#:30:#:;107#E10=:/51.#2?3.#0976:3#6./06#;6#

4H/1B;:7=6#410/54:;#JK410/6P-#6/#;6#4H/1B;:7=6#B01E1.2A#>35#76#E:5/#6.C501.#M#3=#26#;:#
=6=G0:.6#B;:7=5>36F#
):.7#;6#B1;6#C9I9/:/5EA#;67#=5401/3G3;67#71./#B:0:;;<;67#6/#1056./67#2:.7#;:#25064/51.#26#
01/:/51.#410/54:;6#JR^a-F#&6//6#=576#6.#B;:46#76#E:5/#7137#;6#41./0L;6#23#46./051;6#:BB10/9#B:0#
;6#7B60=:/1V1W26F#%.#B0976.46#26#41;48545.6A#;:#01/:/51.#410/54:;6#67/#B60/30G96F##
&6//6#01/:/51.#410/54:;6#6./0:5.6#;14:;6=6./#;6#29B;:46=6./#23#B5I=6./#N#5;#26746.2#:3O
2677137#26#;?9>3:/630#2?3.#4L/9#26#;?Q3E#6/#06=1./6#:3#BL;6#:.5=:;#26#;?:3/06#4L/9F##
Z6;1.#;67#67B<467A#5;#76#E10=6#3.#croissant)dépigmenté#23#4L/9#1BB179#:3#B15./#2?6./096#23#
7B60=:/1V1W26F##
!?:.:;H76#=:401741B5>36#23#29C6;1BB6=6./#3;/905630#26#;?Q3E#:#9/:G;5#>36#;:#09I51.#23#
401577:./##29B5I=6./9#41W.4526#:C64#;:#E3/306#09I51.#2107:;6#26#;?6=G0H1.F#
%.#=6.:./#3.6#6PB9056.46#26#;?500:25:/51.#26#;?Q3E#B:0#267#3;/0:C51;6/7A#23#4L/9#23#B1;6#
C9I9/:/5EA#46;:#:#41.235/##@#;?1G/6./51.#2?3.#6=G0H1.#:/HB5>36#29B130C3#26#7/034/306#2107:;6##
J481026A#71=5/6-#FY.#6=G0H1.#500:259#29B130C36#26#7/034/306#2107:;6#67/#25/#embryon)
centralisé.)
&67#1G760C:/51.7#71./#E1.2:=6./:;67A#4:0#6;;67#75I.5E56./#>36#;:#01/:/51.#410/54:;6#2505I6#;:#
=576#6.#B;:46#23#B;:.#26#7H=9/056#G5;:/90:;6#26#;?6=G0H1.#6/#2?3.#.13C6;#:P6#21071#C6./0:;F#
G- 06257/05G3/51.#267#29/60=5.:./7#4H/1B;:7=5>367##
):.7#;6#B1;6#C9I9/:/5EA#1.#:#3.#460/:5.#.1=G06#26#29/60=5.:./7#4H/1B;:7=5>367F#
#!6#=13C6=6./#23#410/6P#C:#6.I6.2060#;:#06257/05G3/51.#26#467#29/60=5.:./7#N#"U(=A#bc,A#
)Z`A#b6I*F#
"C:./#;:#01/:/51.#23#410/6PA#;:#B01/95.6#)Z`#67/#79>367/096#2:.7#;67#C97543;67#23#410/6P#
C9I9/:/5EF##
"3#=1=6./#26#;:#01/:/51.#410/54:;6A#5;#:#9/9#=1./09#>36#467#C97543;67#C1./#76#06/013C60#
/0:.7;1>397#2:.7#;:#E3/306#09I51.#2107:;6#26#;?6=G0H1.F##
Y.6#E157#:005C967A#2:.7#;:#B:0/56#2107:;6A#;67#C97543;67#;5G<06./#;630#41./6.3A#6/#6.#
;?1443006.46#;:#B01/95.6#)Z`F#
U6=:0>36#N#;:#B01/95.6#)Z`#67/#3.6#B01/95.6#4H/1B;:7=5>36#5./60=925:506#2:.7#;:#C156#26#
75I.:;57:/51.#26#B01/95.67#d+(*F#%;;6#67/#;6#73BB0677630#2?:4/5C5/9#26#;:#B01/95.6#cZeR#
JI;H41I<.6#7H./89/:76#X5.:76#R-F#!:#B01/95.6#cZeR#B01C1>36#;:#29I0:2:/51.#2?3.6#:3/06#
B01/95.6#4H/1B;:7=5>36#N#;:#fO4:/9.5.6##
!:#fO4:/9.5.6#67/#3.6#B01/95.6#=3;/5E1.4/51..6;;6#>35#5./60C56./#2:.7#;?6.40:I6#267#
4:28605.67#:3#4H/17>36;6//6A#J4?67/#3.#E:4/630#.34;9:506#26#/0:.7405B/51.-F#
 6
6
1
/
6
100%