File

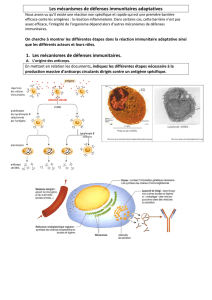
PARTIE III : Enseignement obligatoire (5 points)
Aspects du fonctionnement des centres nerveux
La myasthénie est une maladie neuromusculaire caractérisée par une faiblesse des muscles
squelettiques. Les patients ont des difficultés par exemple à garder leurs paupières levées.
L'impossibilité de mettre en évidence chez les malades des anomalies de structure de
l'innervation du muscle a conduit les médecins à émettre l'hypothèse d'un mauvais
fonctionnement de la jonction neuromusculaire.
A l'aide des documents ci-dessous, éprouvez l'hypothèse émise et proposez un
mécanisme expliquant la myasthénie.
Document 1.
Organisation de la jonction neuromusculaire ou plaque motrice.
Document 2.
Un nerf moteur est stimulé électriquement. Des électrodes posées sur la peau, au niveau d'un
muscle commandé par ce nerf, permettent d'enregistrer les phénomènes électriques globaux
de ce muscle (électromyogramme) lors de la stimulation du nerf.
On enregistre ces phénomènes électriques sur deux sujets, l'un normal, l'autre atteint de
myasthénie.

Document 3.
L'alpha-bungarotoxine, molécule toxique extraite du venin de serpent, possède la propriété de
se fixer sur les récepteurs à acétylcholine. Son injection à une souris saine entraîne des
symptômes analogues à ceux de la myasthénie. Par autoradiographie, on peut localiser l'alpha-
bungarotoxine radioactive sur la membrane d'une cellule musculaire. La figure donne le
résultat de cette localisation chez un individu sain et chez un individu myasthénique.
Document 4.
A l'aide des électrodes réceptrices 1 et 2, on mesure les potentiels dans la zone postsynaptique
de la jonction neuromusculaire. On mesure ainsi le potentiel postsynaptique au niveau de la
plaque motrice (électrode 1) et le potentiel d'action du muscle responsable de la contraction
(électrode 2).
On réalise cette manipulation sur deux jonctions neuromusculaires, d'un individu sain et d'un
individu myasthénique.

PARTIE III : Enseignement de spécialité (5 points)
Aspects du fonctionnement des centres nerveux
La rétine (couche sensorielle de l'œil) possède des cellules spécialisées dans la perception de
la lumière, les bâtonnets. Exposés à la lumière, les bâtonnets génèrent un potentiel de
récepteur à l'origine de potentiels d'action.
En mettant en relation les informations fournies, expliquez comment des échanges
ioniques au niveau des bâtonnets sont à l'origine d'un potentiel de récepteur.
SCHEMA DU MONTAGE EXPERIMENTAL
Deux méthodes d'exploration électrophysiologique permettent d'étudier le fonctionnement
d'un bâtonnet isolé.
Méthode 1 : Mesure du potentiel transmembranaire.
A l'aide d'une électrode transmembranaire on mesure le potentiel de membrane du bâtonnet.

Méthode 2 : Mesure du courant transmembranaire.
On aspire une partie du bâtonnet dans une micropipette. Cette micropipette est utilisée pour
mesurer directement le courant ionique traversant la membrane de la partie aspirée.
Document 1.
Le bâtonnet comporte deux segments sur lesquels on a pu identifier des canaux ioniques
spécifiques au K+, localisés uniquement au niveau du segment interne de cette cellule et des
canaux ioniques spécifiques au Na+ localisés au niveau du segment externe de cette cellule.
Les méthodes 1 et 2 citées dans le montage expérimental ont permis d'établir les phénomènes
ayant lieu à l'obscurité.
Document 2.
On applique à un bâtonnet deux stimulations lumineuses d'intensité croissante (I1 et I2). On
enregistre son potentiel de membrane (A) et les courants transmembranaires (B) à l'aide des
méthodes 1 et 2.

Document 3.
On étudie l'effet de la concentration externe en Na+, notée [Na+]e, sur le fonctionnement de la
cellule. La concentration externe en Na+est de 115 mM, alors que la concentration interne est
de 30 mM. La méthode 2 permet de modifier la solution en contact avec la cellule. On
diminue ainsi la concentration externe en Na+et on stimule le bâtonnet par la lumière
d'intensité I3.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%